

doi: 10.1104/pp.104.041186.
Arabidopsis AtCYP20-2 is a light-regulated cyclophilin-type peptidyl-prolyl cis-trans isomerase associated with the photosynthetic membranes
Affiliations
- PMID: 15084723
- PMCID: PMC419801
- DOI: 10.1104/pp.104.041186
Item in Clipboard
Arabidopsis AtCYP20-2 is a light-regulated cyclophilin-type peptidyl-prolyl cis-trans isomerase associated with the photosynthetic membranes
Plant Physiol.
2004 Apr.
No abstract available
Figures
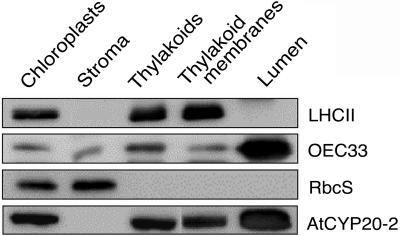
Localization of AtCYP20-2. Western analysis of AtCYP20-2 protein levels in chloroplast fractions (40 μg of protein loaded for AtCYP20-2, 10 μg for OEC33, LHCII, and RbcS). A total of 20 g of Arabidopsis leaf material was ground 7 × 1 s in a blender in 1× grinding buffer (330 mm sorbitol, 20 mm Tricine-KOH, pH 8.4, 5 mm EGTA, 5 mm EDTA, 10 mm NaCO3, 0.1% bovine serum albumin [BSA], and 330 mg/L isoascorbate). The suspension was then filtered through four layers of muslin and centrifuged for 5 min at 1,000g to pellet the crude chloroplast fraction. After resuspension of the chloroplasts in 1× grinding buffer the suspension was layered on top of two 35 mL continuous Percoll gradients. The gradients were generated by 40 min centrifugation at 48,000g of 50% (v/v) Percoll/2× grinding buffer (660 mm sorbitol, 40 mm tricine-KOH, pH 8.4, 10 mm EGTA, 10 mm EDTA, 20 mm NaCO3, 0.2% BSA, and 660 mg/L isoascorbate). The gradient was also supplemented with 132 mg/L (final concentration) of gluthathione. The loaded gradients were centrifuged for 15 min at 3,000g in a swing-out rotor with no brake. The lower band containing intact chloroplasts was collected, washed once in 1× grinding buffer, and diluted to 0.5 mg mL−1 chlorophyll. The chlorophyll concentration during the remainder of the preparation was kept at 0.5 mg mL−1. Intact chloroplasts were then lysed by resuspension in 25 mm NH4HCO3 and incubation on ice for 20 min. The lysed chloroplasts were then centrifuged for 90 s at 16,000g, and the supernatant, consisting of the stromal fraction, was collected and the pelleted thylakoid fraction washed once in 25 mm NH4HCO3 before resuspension in 1× grinding buffer. Intact thylakoids were then lysed by passing them twice through a Yeda press (10 MPa), and the thylakoid membrane and lumen fractions separated by 30 min ultracentrifugation at 200,000g. Thylakoid membranes were finally resuspended in 1× grinding buffer. Protein samples were separated by SDS-PAGE using the buffer system of Davis (1964), and the protocol of Sambrook et al. (1989). A 9% stacking gel and a 15% separating gel were used in all cases. Proteins separated by SDS-PAGE were transferred onto nitrocellulose membrane in transfer buffer (25 mm Tris/HCl, pH 8.3; 192 mm Gly, 20% methanol) with a Bio-Rad blotting kit at 30 mA overnight. The membrane was then blocked in TBS (50 mm Tris-buffered saline (20 mm Tris, 0.5 m NaCl), 5% milk powder, 0.5% Tween 20) for 1 h, followed by 1 h in TBS + primary antibody (dilutions varied according to antibody). Blots were washed for 15 min and 3 × 5 min with Tris-buffered saline followed by incubation in a 1:5000 dilution of the secondary antibody (alkaline phosphatase-conjugated goat anti-rabbit IgG, Amersham Biosciences) for 1 h. The blots were washed again as above and immunodetection was carried out with an enhanced chemiluminescence kit (Amersham Biosciences).
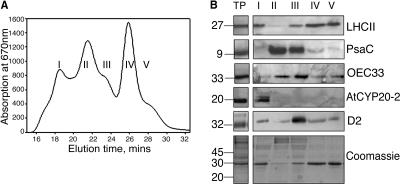
A, Absorbance at 670 nm of fractions obtained from thylakoid membranes purified from LL-grown Columbia-0 plants. B, Western analysis of fractions enriched for (1) PSII supercomplexes, (2) PSI, (3) PSI + PSII core, (4) trimeric LHCII, and (5) monomeric LHCII (protein equivalent of 2 μg of chlorophyll loaded in each well). TP, total thylakoid preparation. Thylakoids were prepared by homogenizing fresh tissue in ice-cold grinding medium (330 mm sorbitol, 5 mm MgCl2, 10 mm Na4P2O7 (pH 6.5), 2 mm ascorbate) with a polytron. The homogenate was then filtered through four layers of muslin followed by two layers of muslin and one layer of cotton wool. The filtrate was centrifuged for 10 min at 5,000g, and the chloroplast enriched pellet resuspended in wash buffer (330 mm sorbitol, 10 mm MES, pH 6.5), followed by a further 10 min centrifugation at 5,000g. The pellet was then resuspended in 5 mm MgCl2 for 30 s to lyse any remaining intact chloroplasts, followed by an equal volume of osmoticum (660 mm sorbitol, 20 mm KCl, 2 mm EDTA and 100 mm HEPES, pH 6.5) to break open the stacked grana. Thylakoid membranes were freshly prepared from 4- to 5-week old Arabidopsis plants as described above with a final resuspension in 20 mm Bis-Tris (pH 6.5), 5 mm MgCl2. Thylakoid samples were diluted to a final chlorophyll concentration of 1.4 mg mL−1 chlorophyll and solubilized by addition of n-dodecyl α-d -maltoside to a final concentration of 0.7% to 0.85%. Samples were vortexed thoroughly for 1 min and centrifuged for 1 min at 16,000g. The supernatant was then filtered through a 0.45-μm nylon filter and subjected to gel filtration chromatography using a Superdex 200 HR 10/30 column in an Amersham-Pharmacia ÄKTApurifier system using the same buffer and similar conditions as before (Yakushevska et al., 2001; Ruban et al., 2003).
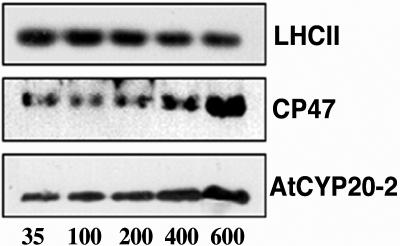
Thylakoid protein extracts from plants grown in steady-state light environments ranging from 35 to 600 μmol m−2 s−1 (inset, representative of 4 experiments) (protein equivalent of 2 μg of chlorophyll/well). Each blot is representative of four separate experiments. Plants were grown under short days (8 h light, 20°C; 16 h dark, 15°C); growth light was provided by color 84 fluorescent tubes (Philips TLD 35 W, or Philips PL-l 55 W heat-filtered through 2 mm polyacetate). Photon fluence rates were measured at the leaf surface using a Skye Instruments (Llandrindod Wells, UK) SKP200 PAR detector.
Comment in
-
Introducing immunophilins. From organ transplantation to plant biology.Plant Physiol. 2004 Apr;134(4):1241-3. doi: 10.1104/pp.103.900108. Plant Physiol. 2004. PMID: 15084722 Free PMC article. No abstract available.
References
-
- Anderson JM, Chow WS, Goodchild DJ (1988) Thylakoid membrane organisation in sun/shade acclimation. Aust J Plant Physiol 15: 11–26
-
- Bailey S, Walters RG, Jansson S, Horton P (2001) Acclimation of Arabidopsis thaliana to the light environment: the existence of separate low light and high light responses. Planta 213: 794–801 - PubMed
-
- Boekema EJ, van Roon H, Calkoen F, Bass R, Dekker JP (1999. a) Multiple types of association of photosystem II and its light-harvesting antenna in partially solubilized photosystem II membranes. Biochemistry 38: 2233–2239 - PubMed
-
- Boekema EJ, van Roon H, Van Breemen JFL, Dekker JP (1999. b) Supramolecular organization of photosystem II and its light-harvesting antenna in partially solubilized photosystem II membranes. Eur J Biochem 266: 444–452 - PubMed
-
- Boekema EJ, Van Breemen JFL, Van Roon H, Dekker JP (2000) Arrangement of photosystem II supercomplexes in crystalline macrodomains within the thylakoid membrane of green plant chloroplasts. J Mol Biol 301: 1123–1133 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
