

Nkx2-5 mutation causes anatomic hypoplasia of the cardiac conduction system
- PMID: 15085192
- PMCID: PMC385399
- DOI: 10.1172/JCI19846
Nkx2-5 mutation causes anatomic hypoplasia of the cardiac conduction system
Abstract
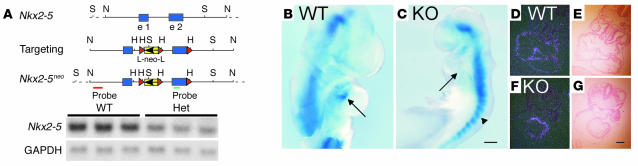
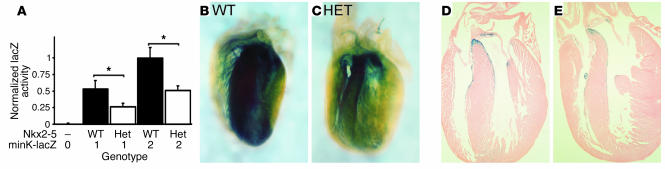
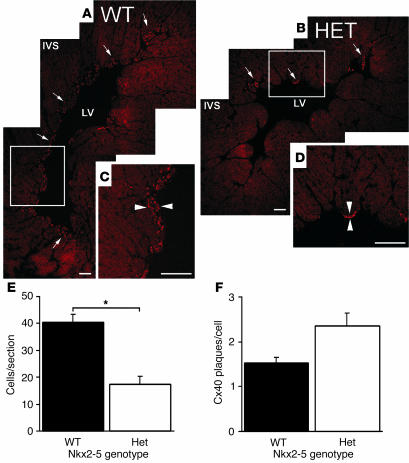
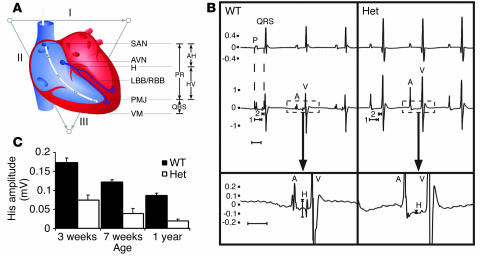
Heterozygous mutations of the cardiac transcription factor Nkx2-5 cause atrioventricular conduction defects in humans by unknown mechanisms. We show in KO mice that the number of cells in the cardiac conduction system is directly related to Nkx2-5 gene dosage. Null mutant embryos appear to lack the primordium of the atrioventricular node. In Nkx2-5 haploinsufficiency, the conduction system has half the normal number of cells. In addition, an entire population of connexin40(-)/connexin45(+) cells is missing in the atrioventricular node of Nkx2-5 heterozygous KO mice. Specific functional defects associated with Nkx2-5 loss of function can be attributed to hypoplastic development of the relevant structures in the conduction system. Surprisingly, the cellular expression of connexin40, the major gap junction isoform of Purkinje fibers and a putative Nkx2-5 target, is unaffected, consistent with normal conduction times through the His-Purkinje system measured in vivo. Postnatal conduction defects in Nkx2-5 mutation may result at least in part from a defect in the genetic program that governs the recruitment or retention of embryonic cardiac myocytes in the conduction system.
Figures

References
-
- Wang, G.F., and Stockdale, F.E. 1999. Chamber-specific gene expression and regulation during heart development. In Heart development. R.P. Harvey and N. Rosenthal, editors. Academic Press. San Diego, California, USA. 357–369.
-
- Cheng G, et al. Development of the cardiac conduction system involves recruitment within a multipotent cardiomyogenic lineage. Development. 1999;126:5041–5049. - PubMed
-
- Gourdie RG, Mima T, Thompson RP, Mikawa T. Terminal diversification of the myocyte lineage generates Purkinje fibers of the cardiac conduction system. Development. 1995;121:1423–1431. - PubMed
-
- Sedmera D, et al. Spatiotemporal pattern of commitment to slowed proliferation in the embryonic mouse heart indicates progressive differentiation of the cardiac conduction system. Anat. Rec. 2003;274A:773–777. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
