

Targeted deletion of BMK1/ERK5 in adult mice perturbs vascular integrity and leads to endothelial failure
- PMID: 15085193
- PMCID: PMC385403
- DOI: 10.1172/JCI19890
Targeted deletion of BMK1/ERK5 in adult mice perturbs vascular integrity and leads to endothelial failure
Abstract
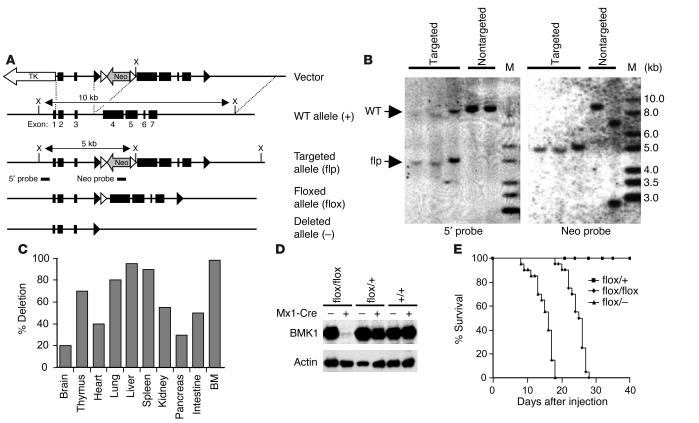
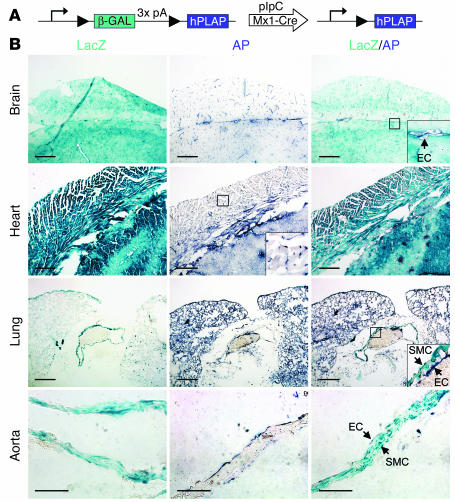
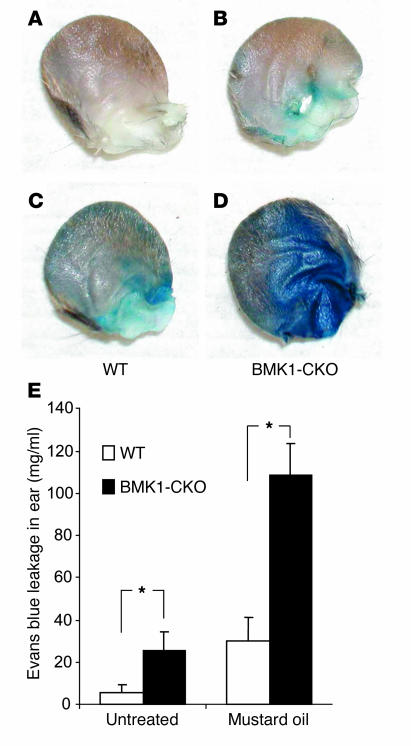
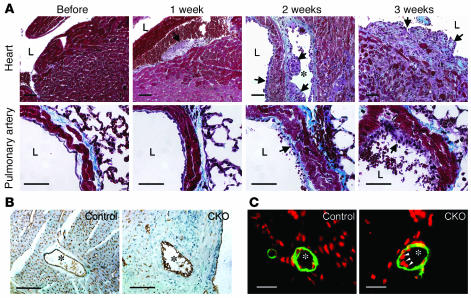
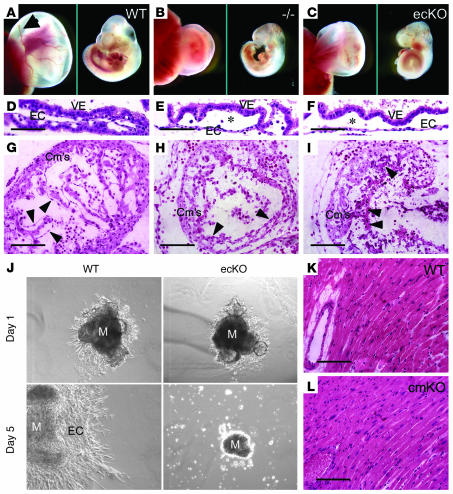
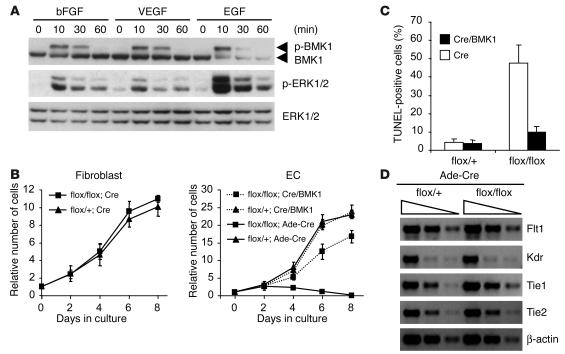
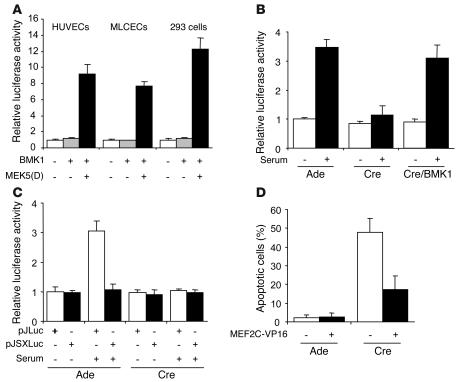
Big mitogen-activated protein kinase 1 (BMK1), also known as ERK5, is a member of the MAPK family. Genetic ablation of BMK1 in mice leads to embryonic lethality, precluding the exploration of pathophysiological roles of BMK1 in adult mice. We generated a BMK1 conditional mutation in mice in which disruption of the BMK1 gene is under the control of the inducible Mx1-Cre transgene. Ablation of BMK1 in adult mice led to lethality within 2-4 weeks after the induction of Cre recombinase. Physiological analysis showed that the blood vessels became abnormally leaky after deletion of the BMK1 gene. Histological analysis revealed that, after BMK1 ablation, hemorrhages occurred in multiple organs in which endothelial cells lining the blood vessels became round, irregularly aligned, and, eventually, apoptotic. In vitro removal of BMK1 protein also led to the death of endothelial cells partially due to the deregulation of transcriptional factor MEF2C, which is a direct substrate of BMK1. Additionally, endothelial-specific BMK1-KO leads to cardiovascular defects identical to that of global BMK1-KO mutants, whereas, surprisingly, mice lacking BMK1 in cardiomyocytes developed to term without any apparent defects. Taken together, the data provide direct genetic evidence that the BMK1 pathway is critical for endothelial function and for maintaining blood vessel integrity.
Figures

Comment in
-
Undermining the endothelium by ablation of MAPK-MEF2 signaling.J Clin Invest. 2004 Apr;113(8):1110-2. doi: 10.1172/JCI21497. J Clin Invest. 2004. PMID: 15085188 Free PMC article.
References
-
- Chao TH, Hayashi M, Tapping RI, Kato Y, Lee JD. MEKK3 directly regulates MEK5 activity as part of the big mitogen-activated protein kinase 1 (BMK1) signaling pathway. J. Biol. Chem. 1999;274:36035–36038. - PubMed
-
- English JM, Vanderbilt CA, Xu S, Marcus S, Cobb MH. Isolation of MEK5 and differential expression of alternatively spliced forms. J. Biol. Chem. 1995;270:28897–28902. - PubMed
-
- Zhou G, Bao ZQ, Dixon JE. Components of a new human protein kinase signal transduction pathway. J. Biol. Chem. 1995;270:12665–12669. - PubMed
-
- Kato Y, et al. Bmk1/Erk5 is required for cell proliferation induced by epidermal growth factor. Nature. 1998;395:713–716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
