

Overexpression of Insig-1 in the livers of transgenic mice inhibits SREBP processing and reduces insulin-stimulated lipogenesis
- PMID: 15085196
- PMCID: PMC385408
- DOI: 10.1172/JCI20978
Overexpression of Insig-1 in the livers of transgenic mice inhibits SREBP processing and reduces insulin-stimulated lipogenesis
Abstract
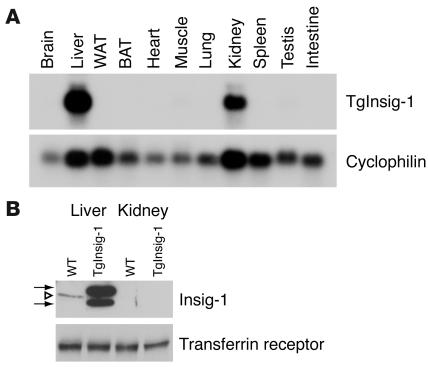
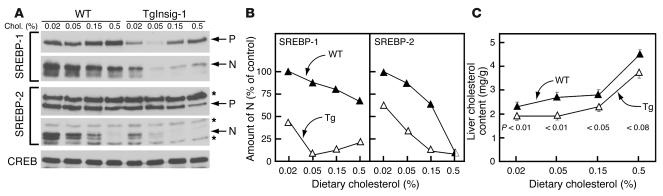
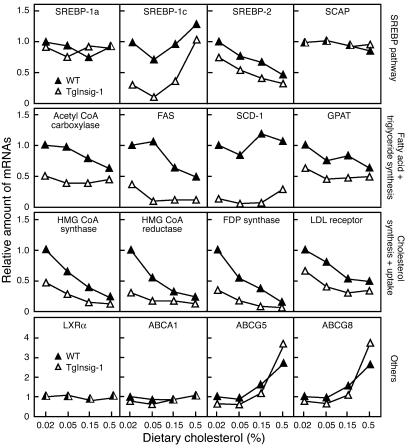
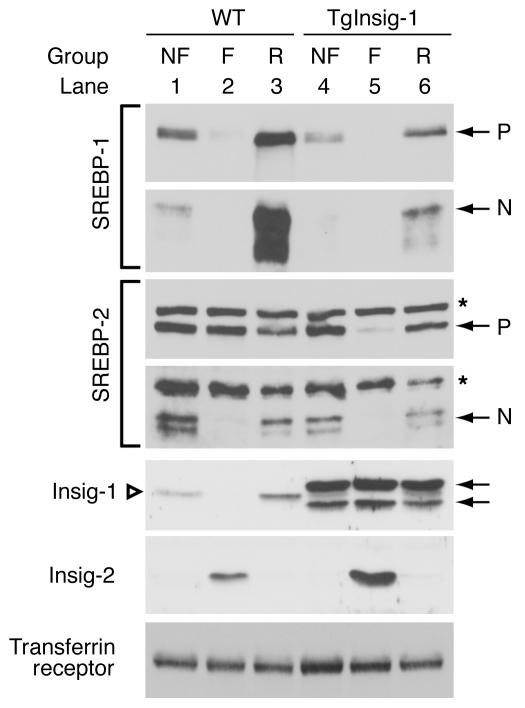
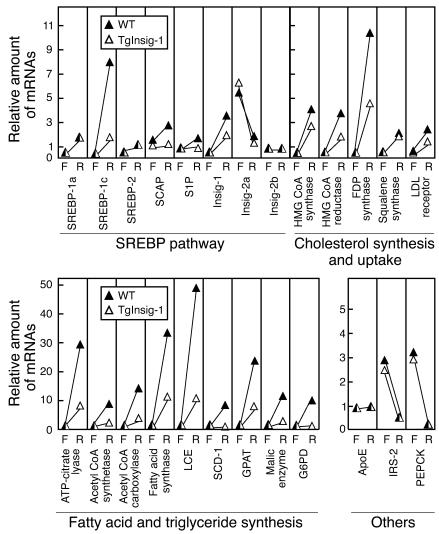
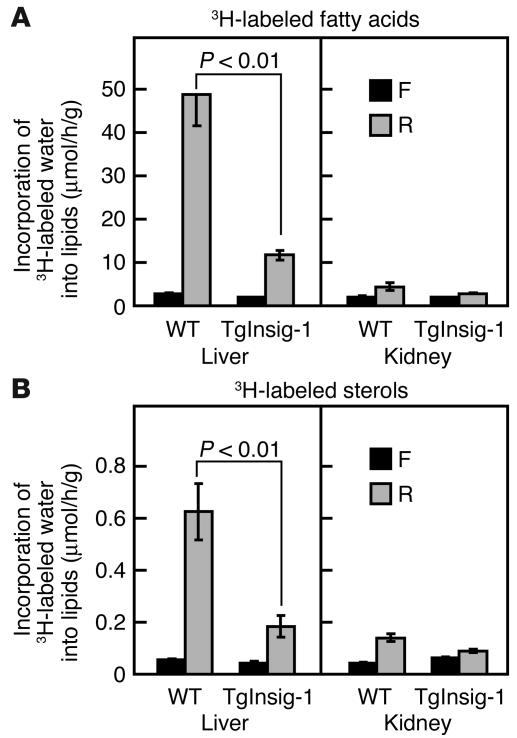
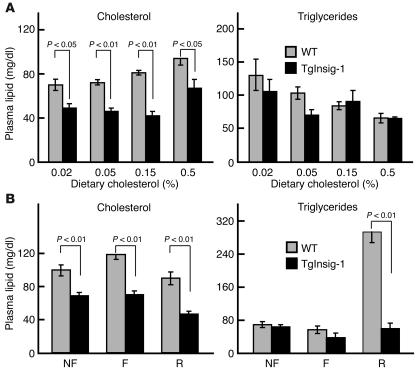
In the current studies we generated transgenic mice that overexpress human Insig-1 in the liver under a constitutive promoter. In cultured cells Insig-1 and Insig-2 have been shown to block lipid synthesis in a cholesterol-dependent fashion by inhibiting proteolytic processing of sterol regulatory element-binding proteins (SREBPs), membrane-bound transcription factors that activate lipid synthesis. Insig's exert this action in the ER by binding SREBP cleavage-activating protein (SCAP) and preventing it from escorting SREBPs to the Golgi apparatus where the SREBPs are processed to their active forms. In the livers of Insig-1 transgenic mice, the content of all nuclear SREBPs (nSREBPs) was reduced and declined further upon feeding of dietary cholesterol. The nuclear content of the insulin-induced SREBP isoform, SREBP-1c, failed to increase to a normal extent upon refeeding on a high-carbohydrate diet. The nSREBP deficiency produced a marked reduction in the levels of mRNAs encoding enzymes required for synthesis of cholesterol, fatty acids, and triglycerides. Plasma cholesterol levels were strongly reduced, and plasma triglycerides did not exhibit their normal rise after refeeding. These results provide in vivo support for the hypothesis that nSREBPs are essential for high levels of lipid synthesis in the liver and indicate that Insig's modulate nSREBP levels by binding and retaining SCAP in the ER.
Figures
Comment in
-
Insig: a significant integrator of nutrient and hormonal signals.J Clin Invest. 2004 Apr;113(8):1112-4. doi: 10.1172/JCI21450. J Clin Invest. 2004. PMID: 15085189 Free PMC article.
References
-
- Sever N, Yang T, Brown MS, Goldstein JL, DeBose-Boyd RA. Accelerated degradation of HMG CoA reductase mediated by binding of insig-1 to its sterol-sensing domain. Mol. Cell. 2003;11:25–33. - PubMed
-
- Sever N, et al. Insig-dependent ubiquitination and degradation of mammalian 3-hydroxy-3-methylglutaryl-CoA reductase stimulated by sterols and geranylgeraniol. J. Biol. Chem. 2003;278:52479–52490. - PubMed
-
- Yang T, et al. Crucial step in cholesterol homeostasis: sterols promote binding of SCAP to INSIG-1, a membrane protein that facilitates retention of SREBPs in ER. Cell. 2002;110:489–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
