

Calcium channels are involved in calcium oxalate crystal formation in specialized cells of Pistia stratiotes L
- PMID: 15087302
- PMCID: PMC4242297
- DOI: 10.1093/aob/mch092
Calcium channels are involved in calcium oxalate crystal formation in specialized cells of Pistia stratiotes L
Abstract
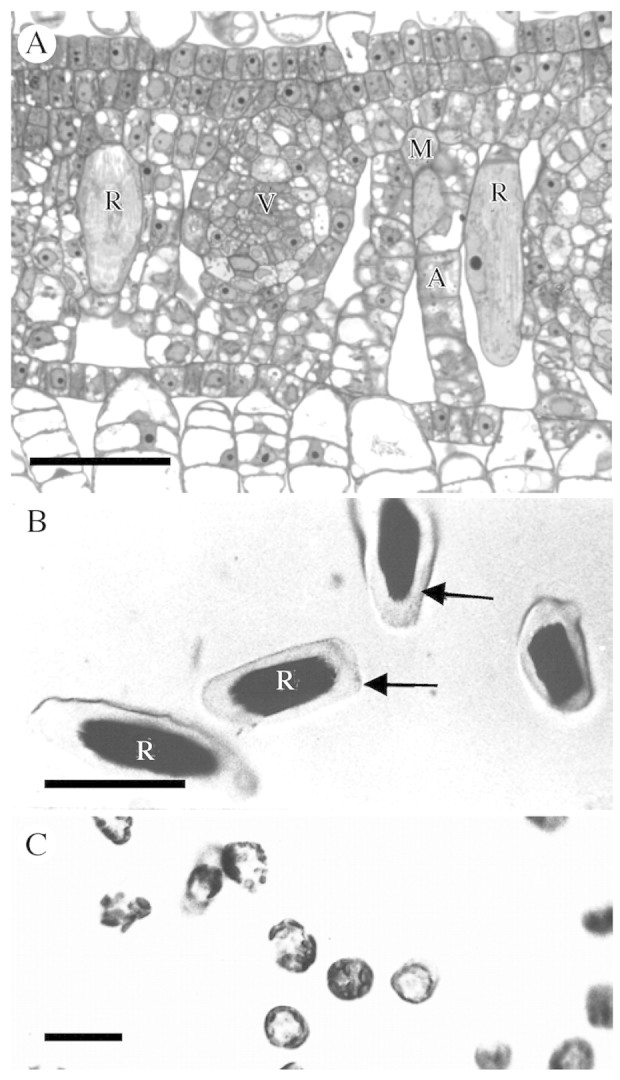
Background and aims: Pistia stratiotes produces large amounts of calcium (Ca) oxalate crystals in specialized cells called crystal idioblasts. The potential involvement of Ca(2+) channels in Ca oxalate crystal formation by crystal idioblasts was investigated.
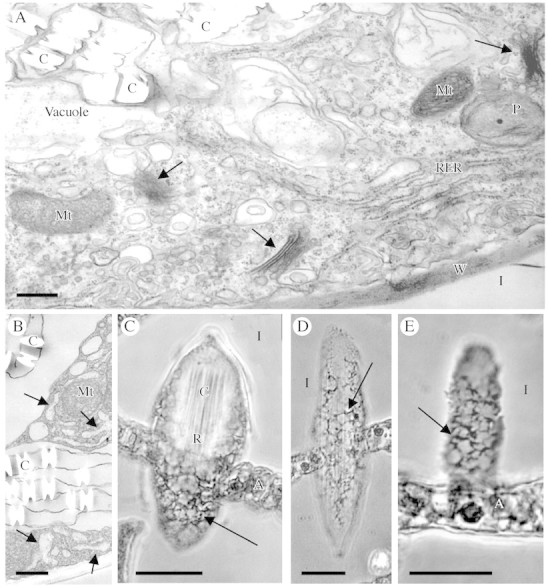
Methods: Anatomical, ultrastructural and physiological analyses were used on plants, fresh or fixed tissues, or protoplasts. Ca(2+) uptake by protoplasts was measured with (45)Ca(2+), and the effect of Ca(2+) channel blockers studied in intact plants. Labelled Ca(2+) channel blockers and a channel protein antibody were used to determine if Ca(2+) channels were associated with crystal idioblasts.
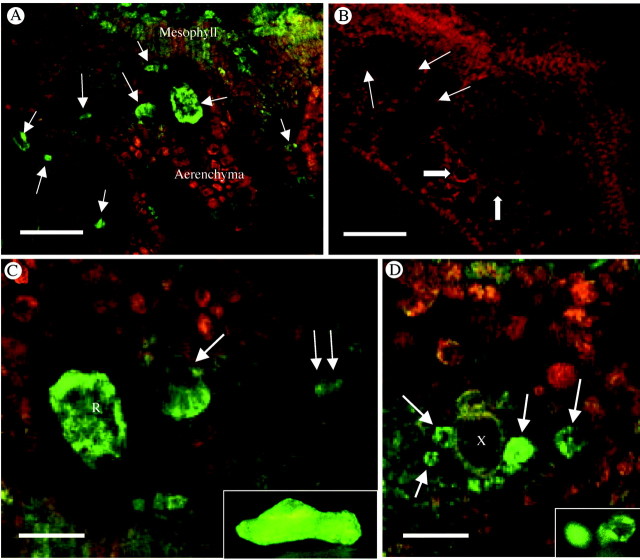
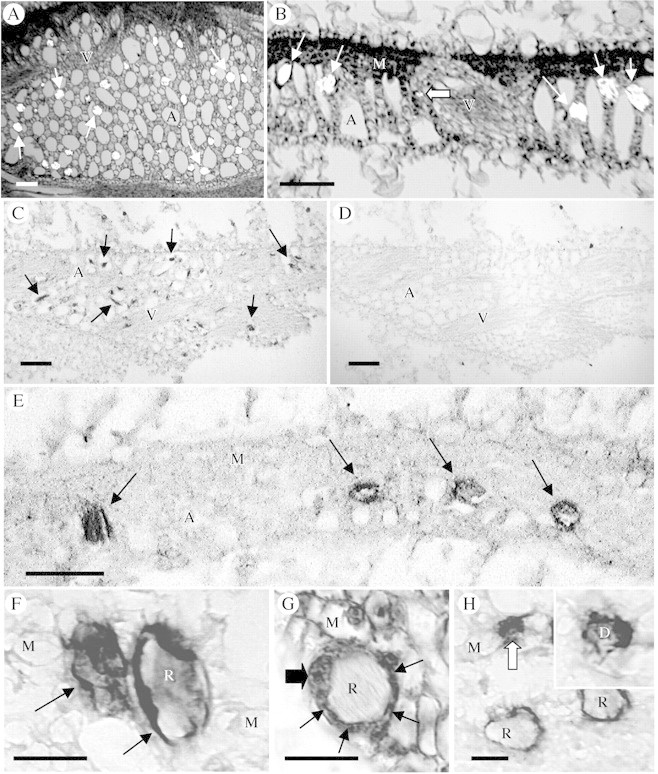
Key results: (45)Ca(2+) uptake was more than two orders of magnitude greater for crystal idioblast protoplasts than mesophyll protoplasts, and idioblast number increased when medium Ca was increased. Plants grown on media containing 1-50 microM of the Ca(2+) channel blockers, isradipine, nifedipine or fluspirilene, showed almost complete inhibition of crystal formation. When fresh tissue sections were treated with the fluorescent dihydropyridine-type Ca(2+) channel blocker, DM-Bodipy-DHP, crystal idioblasts were intensely labelled compared with surrounding mesophyll, and the label appeared to be associated with the plasma membrane and the endoplasmic reticulum, which is shown to be abundant in idioblasts. An antibody to a mammalian Ca(2+) channel alpha1 subunit recognized a single band in a microsomal protein fraction but not soluble protein fraction on western blots, and it selectively and heavily labelled developing crystal idioblasts in tissue sections.
Conclusions: The results demonstrate that Ca oxalate crystal idioblasts are enriched, relative to mesophyll cells, in dihydropyridine-type Ca(2+) channels and that the activity of these channels is important to transport and accumulation of Ca(2+) required for crystal formation.
Figures


References
-
- AndrejauskasE, Hertel R, Marme D.1985. Specific binding of the calcium antagonist [3H]verapamil to membrane fractions from plants. Journal of Biological Chemistry 260: 5411–5414. - PubMed
-
- ArnottHJ.1982. Three systems of biomineralization in plants with comments on the associated organic matrix. In: Nancollas GH, ed. Biological mineralization and demineralization. Berlin: Springer‐Verlag, 199–218.
-
- ArnottHJ, Pautard FGE.1970. Calcification in plants. In: Schraer H, ed. Biological calcification. Cellular and molecular aspects. New York: Appleton‐Century‐Crofts, 375–446.
-
- BhatlaSC, Kiessling J, Reski R.2002. Observation of polarity induction by cytochemical localization of phenylalkylamine‐binding sites in regenerating protoplasts of the moss Physcomitrella patens Protoplasma 219: 99–105. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
