

Genetic re-engineering of Saccharomyces cerevisiae RAD51 leads to a significant increase in the frequency of gene repair in vivo
- PMID: 15087488
- PMCID: PMC407811
- DOI: 10.1093/nar/gkh506
Genetic re-engineering of Saccharomyces cerevisiae RAD51 leads to a significant increase in the frequency of gene repair in vivo
Abstract
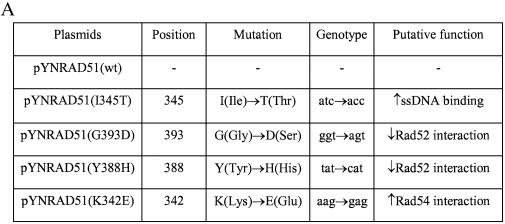
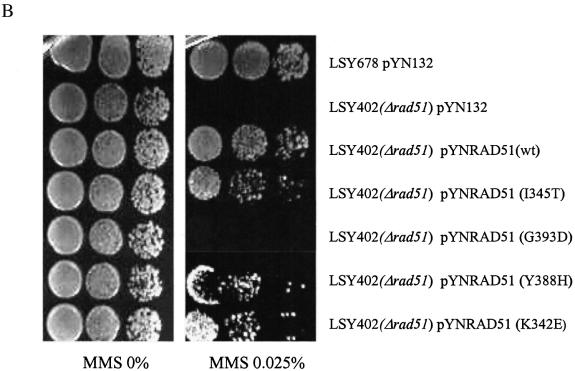
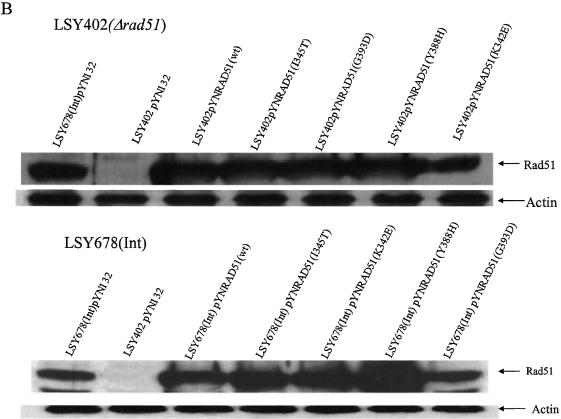
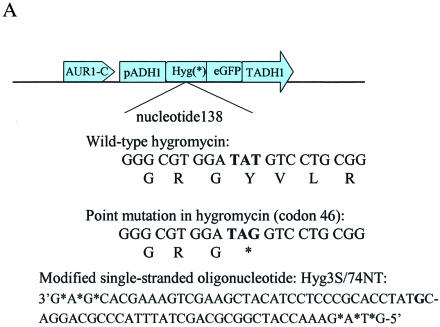
Oligonucleotides can be used to direct the alteration of single nucleotides in chromosomal genes in yeast. Rad51 protein appears to play a central role in catalyzing the reaction, most likely through its DNA pairing function. Here, we re-engineer the RAD51 gene in order to produce proteins bearing altered levels of known activities. Overexpression of wild-type ScRAD51 elevates the correction of an integrated, mutant hygromycin resistance gene approximately 3-fold. Overexpression of an altered RAD51 gene, which encodes a protein that has a higher affinity for ScRad54, enhances the targeting frequency nearly 100-fold. Another mutation which increases the affinity of Rad51 for DNA was also found to increase gene repair when overexpressed in the cell. Other mutations in the Rad51 protein, such as one that reduces interaction with Rad52, has little or no effect on the frequency of gene repair. These data provide the first evidence that the Rad51 protein can be modified so as to increase the frequency of gene repair in yeast.
Figures

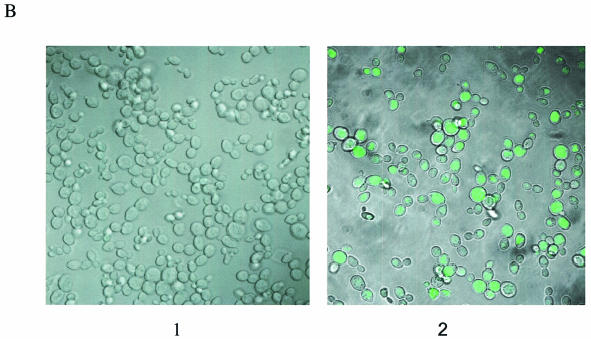
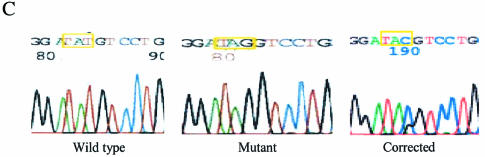
References
-
- Shinohara A., Ogawa,H. and Ogawa,T. (1992) Rad51 protein involved in repair and recombination in S.cerevisiae is a RecA-like protein. Cell, 69, 457–470. - PubMed
-
- Shinohara A., Gasior,S., Ogawa,T., Kleckner,N. and Bishop,D.K. (1997) Saccharomyces cerevisiae recA homologues RAD51 and DMC1 have both distinct and overlapping roles in meiotic recombination. Genes Cells, 2, 615–629. - PubMed
-
- Sugiyama T., Zaitseva,E.M. and Kowalczykowski,S.C. (1997) A single-stranded DNA-binding protein is needed for efficient presynaptic complex formation by the Saccharomyces cerevisiae Rad51 protein. J. Biol. Chem., 272, 7940–7945. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
