

Dominant inhibitory adipocyte-specific secretory factor (ADSF)/resistin enhances adipogenesis and improves insulin sensitivity
- PMID: 15090646
- PMCID: PMC404122
- DOI: 10.1073/pnas.0305905101
Dominant inhibitory adipocyte-specific secretory factor (ADSF)/resistin enhances adipogenesis and improves insulin sensitivity
Abstract
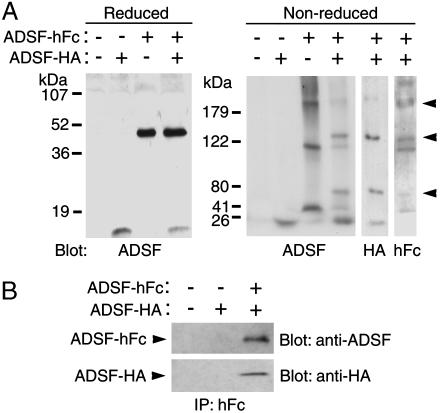
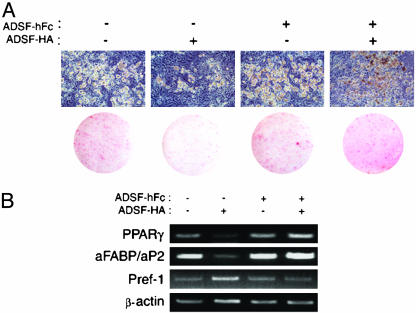
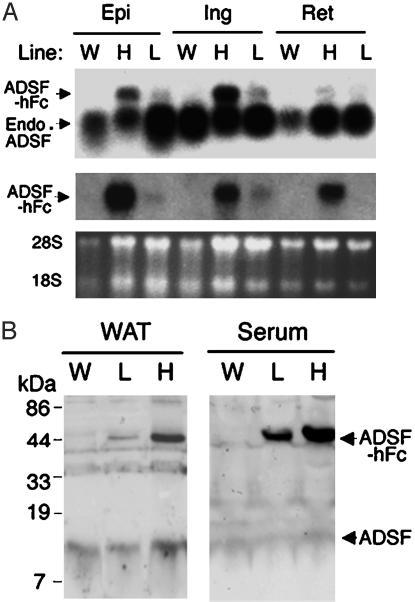
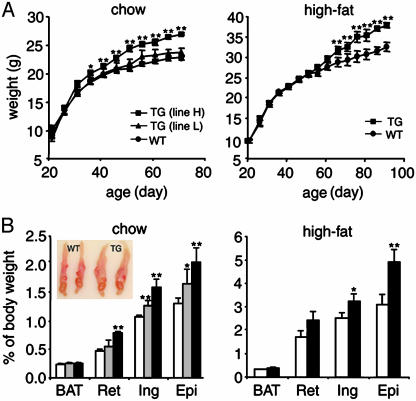
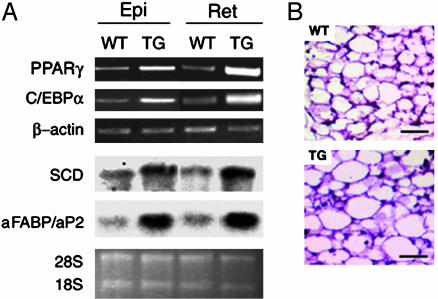
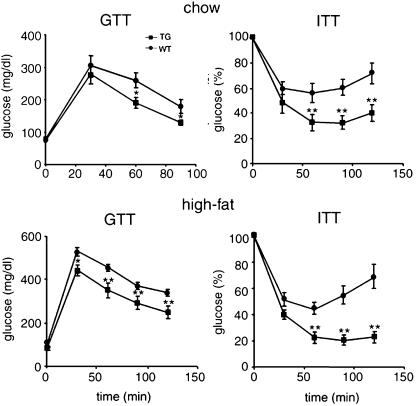
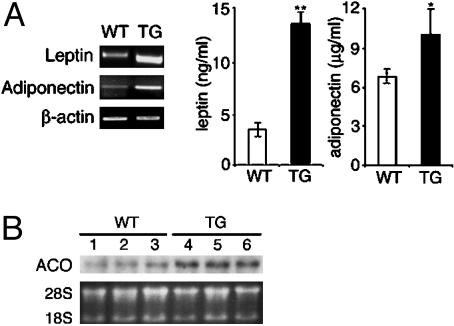
Adipocyte-specific secretory factor (ADSF)/resistin is a small cysteine-rich protein secreted from adipose tissue that belongs to a gene family found in inflammatory zone (FIZZ) or found in resistin-like molecule (RELM). ADSF has been implicated in modulating adipogenesis and insulin resistance. To examine the long-term function of ADSF in adipogenesis and glucose homeostasis, we constructed an expression vector for a dominant inhibitory form of ADSF by fusing it to the human IgGgamma constant region (hFc). ADSF-hFc not only homodimerizes but heterooligomerizes with ADSF/resistin and prevents ADSF/resistin inhibition of adipocyte differentiation of 3T3-L1 cells in a dominant negative manner. Transgenic mice overexpressing ADSF-hFc in adipose tissue show increased adiposity with elevated expression of adipocyte markers as well as enlarged adipocyte size. This finding clearly demonstrates in vivo the inhibitory role of ADSF in adipogenesis. ADSF-hFc transgenic mice with impaired ADSF function exhibit improved glucose tolerance and insulin sensitivity either on chow or high-fat diets. Because of the enhanced adipocyte differentiation, the ADSF-hFc transgenic mice show increased expression of leptin and adiponectin in adipose tissue. The elevated circulating levels for these adipocyte-derived hormones with decreased plasma triglyceride and free fatty acid levels may account for the improved glucose and insulin tolerance in these transgenic mice.
Figures
References
-
- Klyde, B. J. & Hirsch, J. (1979) J. Lipid Res. 20, 705-715. - PubMed
-
- Gregoire, F. M., Smas, C. M. & Sul, H. S. (1998) Physiol. Rev. 78, 783-809. - PubMed
-
- MacDougald, O. A. & Mandrup, S. (2002) Trends Endocrinol. Metab. 13, 5-11. - PubMed
-
- Zhao, L., Gregoire, F. & Sul, H. S. (2000) J. Biol. Chem. 275, 16845-16850. - PubMed
-
- Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L. & Friedman, J. M. (1994) Nature 372, 425-432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
