

Essential role of Sox9 in the pathway that controls formation of cardiac valves and septa
- PMID: 15096597
- PMCID: PMC404074
- DOI: 10.1073/pnas.0401711101
Essential role of Sox9 in the pathway that controls formation of cardiac valves and septa
Abstract
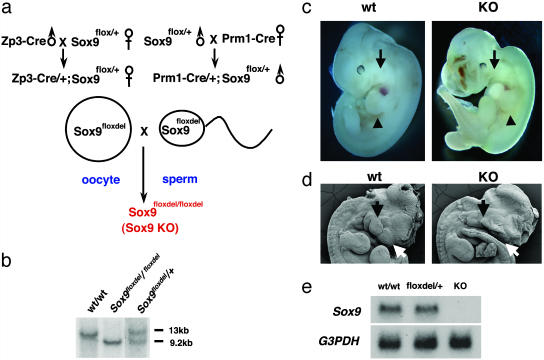
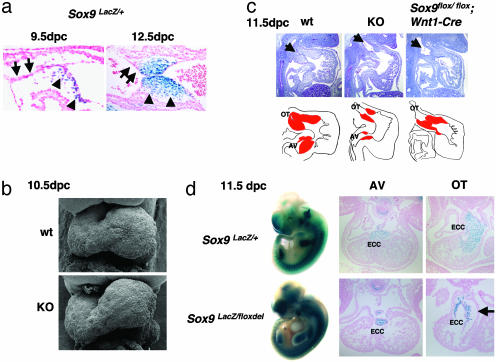
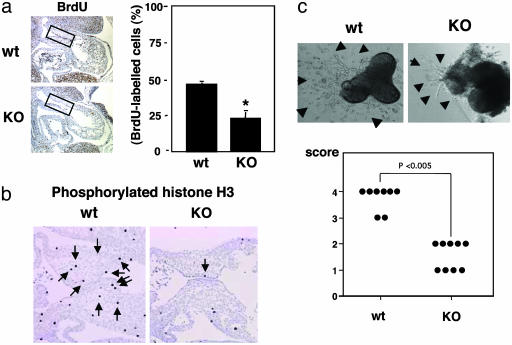
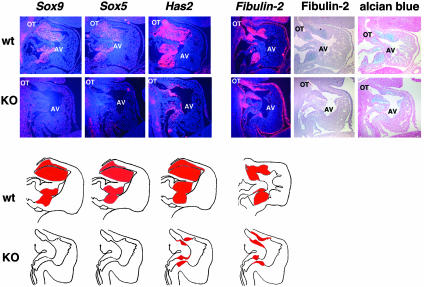
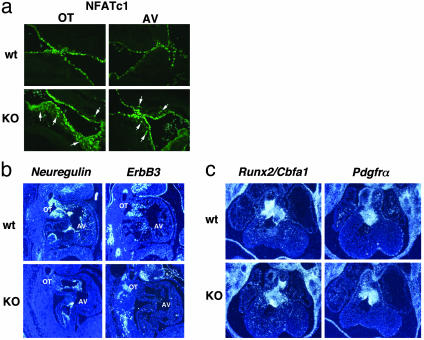
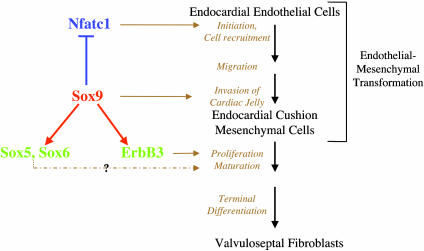
Epithelial-mesenchymal transformation is a critical developmental process reiterated in multiple organs throughout embryogenesis. Formation of endocardial cushions, primordia of valves and septa, is a classic example of epithelial-mesenchymal transformation. Several gene mutations are known to affect cardiac valve formation. Sox9 is activated when endocardial endothelial cells undergo mesenchymal transformation and migrate into an extracellular matrix, called cardiac jelly, to form endocardial cushions. In Sox9-null mutants, endocardial cushions are markedly hypoplastic. In these mutants, Nfatc1 is ectopically expressed and no longer restricted to endothelial cells. Further, Sox9-deficient endocardial mesenchymal cells fail to express ErbB3, which is required for endocardial cushion cell differentiation and proliferation. Our results reveal a succession of molecular steps in the pathway of endocardial cushion development. We propose that loss of Sox9 inhibits epithelial-mesenchymal transformation after delamination and initial migration, but before definitive mesenchymal transformation.
Figures
References
-
- Zhao, Q., Eberspaecher, H., Lefebvre, V. & De Crombrugghe, B. (1997) Dev. Dyn. 209, 377-386. - PubMed
-
- Ng, L. J., Wheatley, S., Muscat, G. E., Conway-Campbell, J., Bowles, J., Wright, E., Bell, D. M., Tam, P. P., Cheah, K. S. & Koopman, P. (1997) Dev. Biol. 183, 108-121. - PubMed
-
- Wagner, T., Wirth, J., Meyer, J., Zabel, B., Held, M., Zimmer, J., Pasantes, J., Bricarelli, F. D., Keutel, J., Hustert, E., et al. (1994) Cell 79, 1111-1120. - PubMed
-
- Foster, J. W., Dominguez-Steglich, M. A., Guioli, S., Kowk, G., Weller, P. A., Stevanovic, M., Weissenbach, J., Mansour, S., Young, I. D., Goodfellow, P. N., et al. (1994) Nature 372, 525-530. - PubMed
-
- Houston, C. S., Opitz, J. M., Spranger, J. W., Macpherson, R. I., Reed, M. H., Gilbert, E. F., Herrmann, J. & Schinzel, A. (1983) Am. J. Med. Genet. 15, 3-28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
