

Human posterior auditory cortex gates novel sounds to consciousness
- PMID: 15096618
- PMCID: PMC404127
- DOI: 10.1073/pnas.0303760101
Human posterior auditory cortex gates novel sounds to consciousness
Abstract
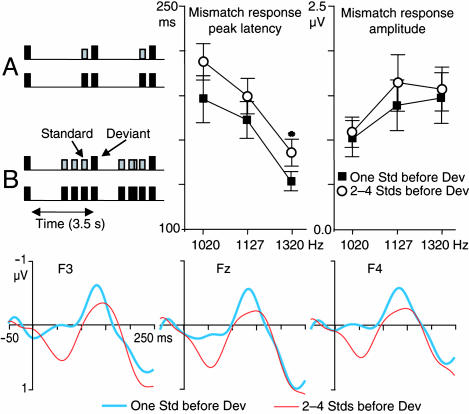
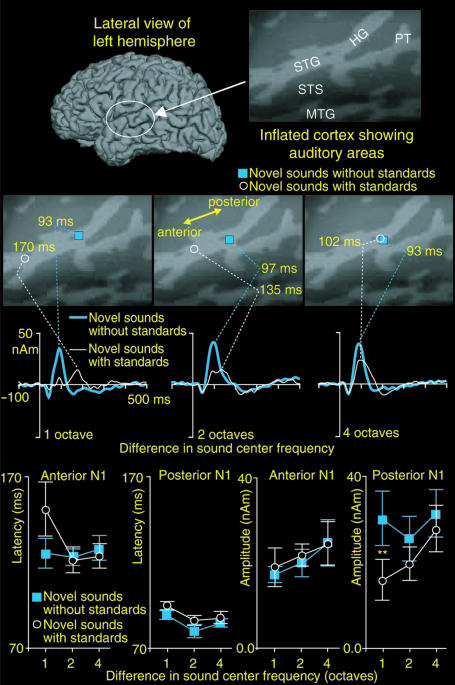
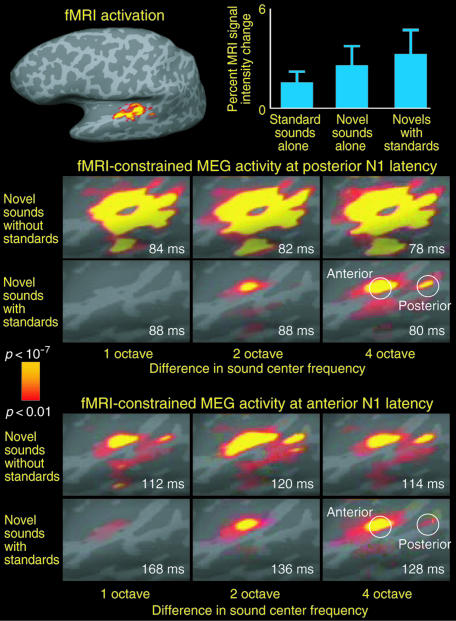
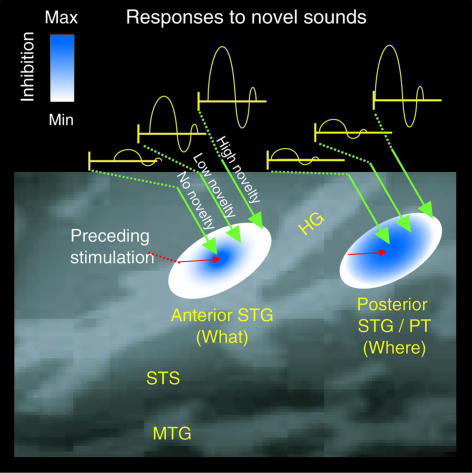
Life or death in hostile environments depends crucially on one's ability to detect and gate novel sounds to awareness, such as that of a twig cracking under the paw of a stalking predator in a noisy jungle. Two distinct auditory cortex processes have been thought to underlie this phenomenon: (i) attenuation of the so-called N1 response with repeated stimulation and (ii) elicitation of a mismatch negativity response (MMN) by changes in repetitive aspects of auditory stimulation. This division has been based on previous studies suggesting that, unlike for the N1, repetitive "standard" stimuli preceding a physically different "novel" stimulus constitute a prerequisite to MMN elicitation, and that the source loci of MMN and N1 are different. Contradicting these findings, our combined electromagnetic, hemodynamic, and psychophysical data indicate that the MMN is generated as a result of differential adaptation of anterior and posterior auditory cortex N1 sources by preceding auditory stimulation. Early ( approximately 85 ms) neural activity within posterior auditory cortex is adapted as sound novelty decreases. This alters the center of gravity of electromagnetic N1 source activity, creating an illusory difference between N1 and MMN source loci when estimated by using equivalent current dipole fits. Further, our electroencephalography data show a robust MMN after a single standard event when the interval between two consecutive novel sounds is kept invariant. Our converging findings suggest that transient adaptation of feature-specific neurons within human posterior auditory cortex filters superfluous sounds from entering one's awareness.
Figures

References
-
- James, W. (1890) Principles of Psychology (Holt, New York).
-
- Hari, R., Kaila, K., Katila, T., Tuomisto, T. & Varpula, T. (1982) Electroencephalogr. Clin. Neurophysiol. 54, 561-569. - PubMed
-
- Tiitinen, H., Alho, K., Huotilainen, M., Ilmoniemi, R. J., Simola, J. & Näätänen, R. (1993) Psychophysiology 30, 537-540. - PubMed
-
- Woldorff, M. G., Hillyard, S. A., Gallen, C. C., Hampson, S. R. & Bloom, F. E. (1998) Psychophysiology 35, 283-292. - PubMed
-
- Näätänen, R. (1992) Attention and Brain Function (Erlbaum, Mahwah, NJ).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
