

Extracellular invertase is an essential component of cytokinin-mediated delay of senescence
- PMID: 15100396
- PMCID: PMC423215
- DOI: 10.1105/tpc.018929
Extracellular invertase is an essential component of cytokinin-mediated delay of senescence
Abstract
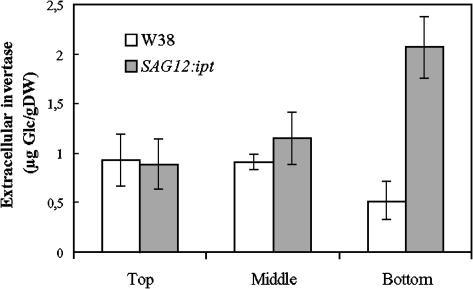
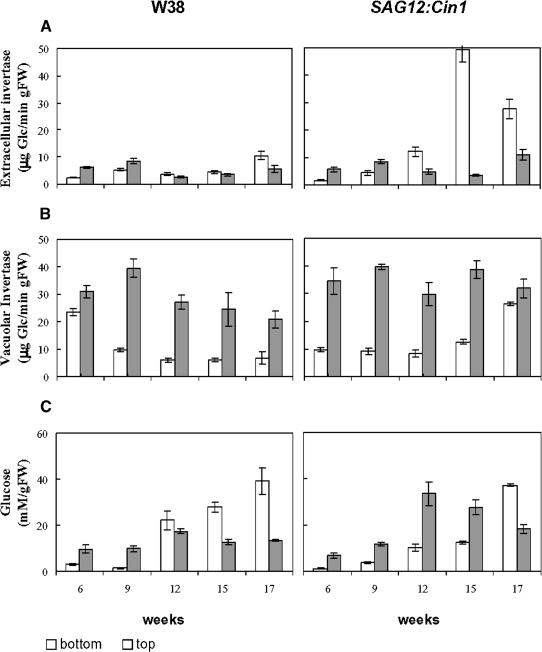
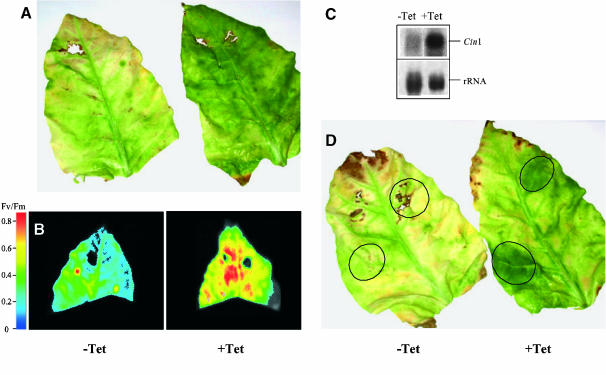
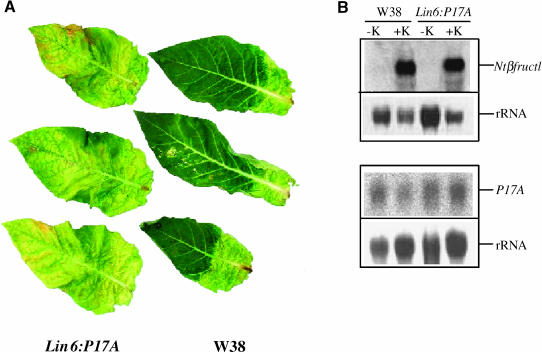
Leaf senescence is the final stage of leaf development in which the nutrients invested in the leaf are remobilized to other parts of the plant. Whereas senescence is accompanied by a decline in leaf cytokinin content, exogenous application of cytokinins or an increase of the endogenous concentration delays senescence and causes nutrient mobilization. The finding that extracellular invertase and hexose transporters, as the functionally linked enzymes of an apolasmic phloem unloading pathway, are coinduced by cytokinins suggested that delay of senescence is mediated via an effect on source-sink relations. This hypothesis was further substantiated in this study by the finding that delay of senescence in transgenic tobacco (Nicotiana tabacum) plants with autoregulated cytokinin production correlates with an elevated extracellular invertase activity. The finding that the expression of an extracellular invertase under control of the senescence-induced SAG12 promoter results in a delay of senescence demonstrates that effect of cytokinins may be substituted by these metabolic enzymes. The observation that an increase in extracellular invertase is sufficient to delay leaf senescence was further verified by a complementing functional approach. Localized induction of an extracellular invertase under control of a chemically inducible promoter resulted in ectopic delay of senescence, resembling the naturally occurring green islands in autumn leaves. To establish a causal relationship between cytokinins and extracellular invertase for the delay of senescence, transgenic plants were generated that allowed inhibition of extracellular invertase in the presence of cytokinins. For this purpose, an invertase inhibitor was expressed under control of a cytokinin-inducible promoter. It has been shown that senescence is not any more delayed by cytokinin when the expression of the invertase inhibitor is elevated. This finding demonstrates that extracellular invertase is required for the delay of senescence by cytokinins and that it is a key element of the underlying molecular mechanism.
Figures

References
-
- Angra, S.R., and Mandahar, C.L. (1993). Involvement of carbohydrates and cytokinins in pathogenicity of Helminthosporium carbonum. Mycopathologia 121, 91–99.
-
- Bradford, M.M. (1976). A rapid and sensitive method for the determination of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. - PubMed
-
- Brenner, M.L., and Cheikh, N. (1995). The role of phytohormones in photosynthate partitioning and seed filling. In Plant Hormones, P.J. Davies, ed (Dordrecht, The Netherlands: Kluwer Academic Press), pp. 649–670.
-
- Buchanan-Wollaston, V. (1997). The molecular biology of leaf senescence. J. Exp. Bot. 48, 181–199.
-
- Chandlee, J.M. (2001). Current molecular understanding of the genetically programmed process of leaf senescence. Physiol. Plant 113, 1–8.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
