

Membrane-associated guanylate kinase-like properties of beta-subunits required for modulation of voltage-dependent Ca2+ channels
- PMID: 15100405
- PMCID: PMC406488
- DOI: 10.1073/pnas.0306665101
Membrane-associated guanylate kinase-like properties of beta-subunits required for modulation of voltage-dependent Ca2+ channels
Abstract
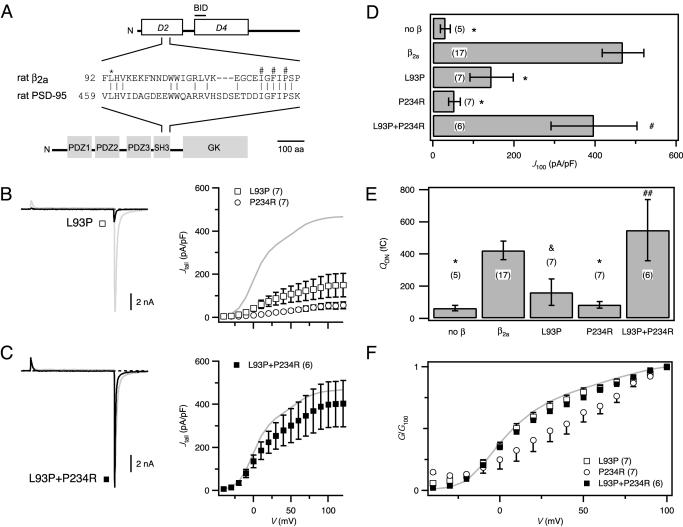
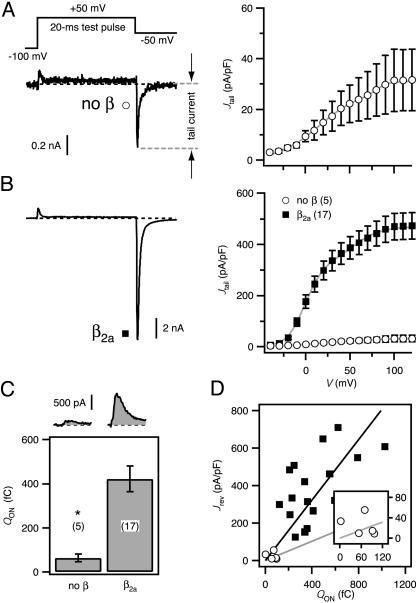
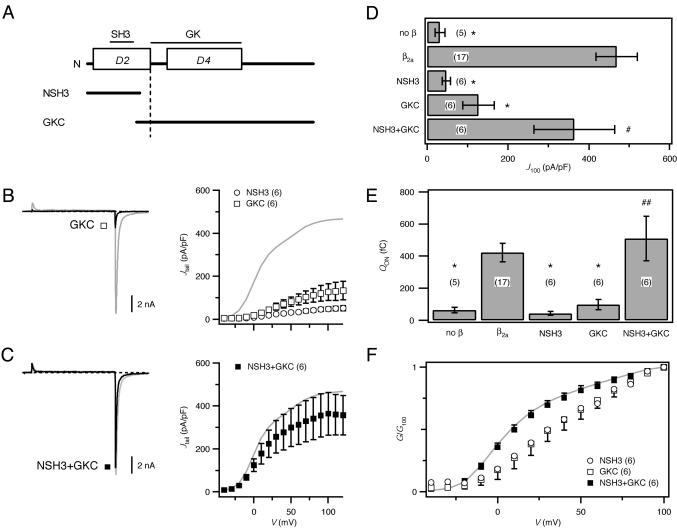
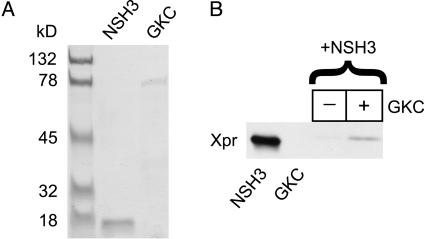
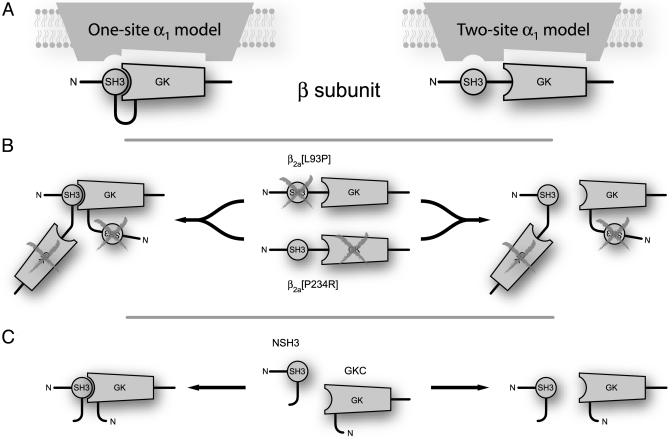
High-voltage-activated Ca2+ channels regulate diverse functions ranging from muscle contraction to synaptic transmission. Association between auxiliary beta- and distinct pore-forming alpha1-subunits is obligatory for forming functional high-voltage-activated Ca2+ channels, yet the structural determinants underlying this interaction remain poorly understood. Recently, homology modeling of Ca(2+)-channel beta1b-subunit identified src homology 3 (SH3) and guanylate kinase (GK) motifs in a tandem arrangement reminiscent of the membrane-associated guanylate kinase (MAGUK) class of scaffolding proteins. However, direct evidence for MAGUK-like properties and their functional implications in beta-subunits is lacking. Here, we show a functional requirement for both SH3 and GK domains in beta2a. Point mutations in either the putative beta2a SH3 or GK domains severely blunted modulation of recombinant L-type channels, showing the importance of both motifs for a functional alpha1-beta interaction. Coexpression of these functionally deficient beta2a-SH3 and GK mutants rescued WT currents, demonstrating trans complementation similar to that observed in MAGUKs. Truncated "hemi-beta2a" subunits, containing either the SH3 or GK domain, were ineffective on their own, but reconstituted WT currents when coexpressed. Moreover, the SH3 and GK domains were found to interact in vitro. These findings reveal MAGUK-like properties in beta-subunits that are critical for alpha1-subunit modulation, revise current models of alpha1-beta association, and predict new physiological dimensions of beta-subunit function.
Figures
Similar articles
-
A CaVbeta SH3/guanylate kinase domain interaction regulates multiple properties of voltage-gated Ca2+ channels.J Gen Physiol. 2005 Oct;126(4):365-77. doi: 10.1085/jgp.200509354. J Gen Physiol. 2005. PMID: 16186563 Free PMC article.
-
The HOOK-domain between the SH3 and the GK domains of Cavbeta subunits contains key determinants controlling calcium channel inactivation.Channels (Austin). 2007 Mar-Apr;1(2):92-101. doi: 10.4161/chan.4145. Epub 2007 Mar 15. Channels (Austin). 2007. PMID: 18690022
-
An intramolecular interaction between Src homology 3 domain and guanylate kinase-like domain required for channel clustering by postsynaptic density-95/SAP90.J Neurosci. 2000 May 15;20(10):3580-7. doi: 10.1523/JNEUROSCI.20-10-03580.2000. J Neurosci. 2000. PMID: 10804199 Free PMC article.
-
[Structure of the calcium channel beta subunit: the place of the beta-interaction domain].Med Sci (Paris). 2005 Mar;21(3):279-83. doi: 10.1051/medsci/2005213279. Med Sci (Paris). 2005. PMID: 15745702 Review. French.
-
The ß subunit of voltage-gated Ca2+ channels.Physiol Rev. 2010 Oct;90(4):1461-506. doi: 10.1152/physrev.00057.2009. Physiol Rev. 2010. PMID: 20959621 Free PMC article. Review.
Cited by
-
Forskolin Regulates L-Type Calcium Channel through Interaction between Actinin 4 and β3 Subunit in Osteoblasts.PLoS One. 2015 Apr 22;10(4):e0124274. doi: 10.1371/journal.pone.0124274. eCollection 2015. PLoS One. 2015. PMID: 25902045 Free PMC article.
-
The secret lives of voltage sensors.J Physiol. 2007 Sep 15;583(Pt 3):813-4. doi: 10.1113/jphysiol.2007.141077. Epub 2007 Aug 9. J Physiol. 2007. PMID: 17690136 Free PMC article. Review. No abstract available.
-
A CaV1.1 Ca2+ channel splice variant with high conductance and voltage-sensitivity alters EC coupling in developing skeletal muscle.Biophys J. 2009 Jan;96(1):35-44. doi: 10.1016/j.bpj.2008.09.027. Biophys J. 2009. PMID: 19134469 Free PMC article.
-
A CaVbeta SH3/guanylate kinase domain interaction regulates multiple properties of voltage-gated Ca2+ channels.J Gen Physiol. 2005 Oct;126(4):365-77. doi: 10.1085/jgp.200509354. J Gen Physiol. 2005. PMID: 16186563 Free PMC article.
-
β subunits of voltage-gated calcium channels in cardiovascular diseases.Front Cardiovasc Med. 2023 Feb 2;10:1119729. doi: 10.3389/fcvm.2023.1119729. eCollection 2023. Front Cardiovasc Med. 2023. PMID: 36818347 Free PMC article. Review.
References
-
- Tsien, R. W. & Tsien, R. Y. (1990) Annu. Rev. Cell Biol. 6, 715-760. - PubMed
-
- Hille, B. (2001) Ion Channels of Excitable Membranes (Sinauer, Sunderland, MA).
-
- Catterall, W. A. (2000) Annu. Rev. Cell Dev. Biol. 16, 521-555. - PubMed
-
- Chien, A. J., Zhao, X., Shirokov, R. E., Puri, T. S., Chang, C. F., Sun, D., Rios, E. & Hosey, M. M. (1995) J. Biol. Chem. 270, 30036-30044. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
