

Tob deficiency superenhances osteoblastic activity after ovariectomy to block estrogen deficiency-induced osteoporosis
- PMID: 15100414
- PMCID: PMC404100
- DOI: 10.1073/pnas.0303093101
Tob deficiency superenhances osteoblastic activity after ovariectomy to block estrogen deficiency-induced osteoporosis
Abstract
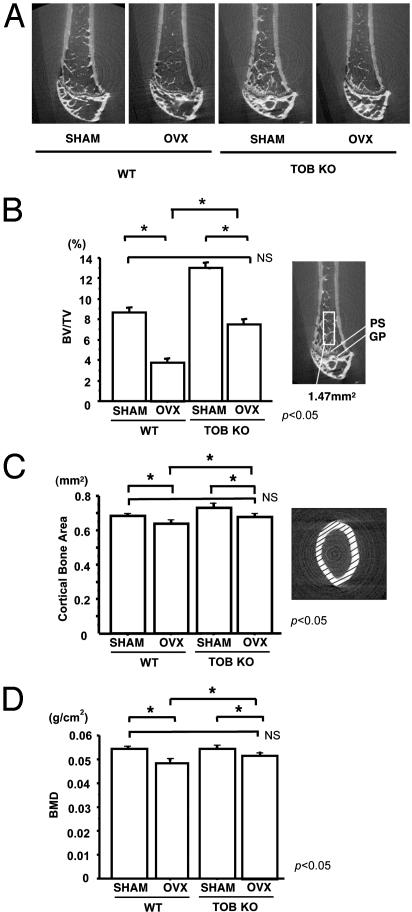
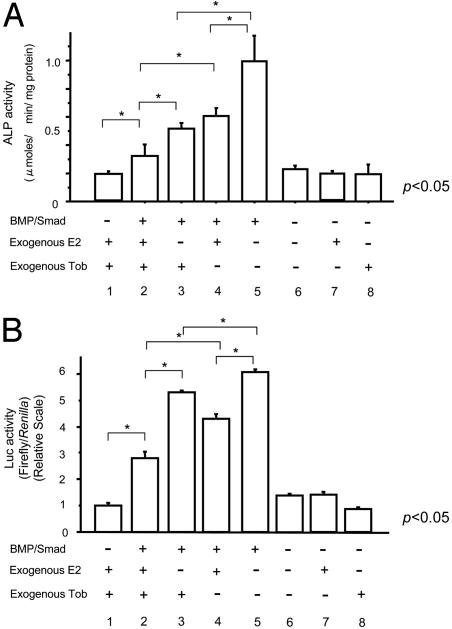
Tob (transducer of erbB2) is a member of antiproliferative family proteins and acts as a bone morphogenic protein inhibitor as well as a suppressor of proliferation in T cells, which have been implicated in postmenopausal bone loss. To determine the effect of Tob deficiency on estrogen deficiency-induced bone loss, we analyzed bone metabolism after ovariectomy or sham operation in Tob-deficient mice. Ovariectomy in WT mice decreased trabecular bone volume and bone mineral density (BMD) as expected. In Tob-deficient mice, ovariectomy reduced bone volume and BMD. However, even after ovariectomy, both trabecular bone volume and BMD levels in Tob-deficient bone were comparable to those in sham-operated WT bones. Bone formation parameters (mineral apposition rate and bone formation rate) in the ovariectomized Tob-deficient mice were significantly higher than those in the ovariectomized WT mice. In contrast, the ovariectomy-induced increase in the bone resorption parameters, osteoclast surface, and osteoclast number was similar between Tob-deficient mice and WT mice. Furthermore, in ex vivo nodule formation assay, ovariectomy-induced enhancement of nodule formation was significantly higher in the bone marrow cells from Tob-deficient mice than in the bone marrow cells from ovariectomized WT mice. Both Tob and estrogen signalings converge at bone morphogenic protein activation of alkaline phosphatase and GCCG-reporter gene expression in osteoblasts, revealing interaction between the two signals. These data indicate that Tob deficiency prevents ovariectomy-induced bone loss through the superenhancement of osteoblastic activities in bone and that this results in further augmentation in the bone formation rate and the mineral apposition rate after ovariectomy in vivo.
Figures





Similar articles
-
Biglycan deficiency interferes with ovariectomy-induced bone loss.J Bone Miner Res. 2003 Dec;18(12):2152-8. doi: 10.1359/jbmr.2003.18.12.2152. J Bone Miner Res. 2003. PMID: 14672350
-
Interleukin-7 influences osteoclast function in vivo but is not a critical factor in ovariectomy-induced bone loss.J Bone Miner Res. 2006 May;21(5):695-702. doi: 10.1359/jbmr.060117. J Bone Miner Res. 2006. PMID: 16734384
-
Inducible nitric oxide synthase mediates bone loss in ovariectomized mice.Endocrinology. 2003 Mar;144(3):1098-107. doi: 10.1210/en.2002-220597. Endocrinology. 2003. PMID: 12586786
-
Cellular and molecular effects of growth hormone and estrogen on human bone cells.APMIS Suppl. 1997;71:1-30. APMIS Suppl. 1997. PMID: 9357492 Review.
-
Immune regulation of bone loss by Th17 cells in oestrogen-deficient osteoporosis.Eur J Clin Invest. 2013 Nov;43(11):1195-202. doi: 10.1111/eci.12158. Epub 2013 Sep 5. Eur J Clin Invest. 2013. PMID: 24033116 Review.
Cited by
-
TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease.Bone Res. 2016 Apr 26;4:16009. doi: 10.1038/boneres.2016.9. eCollection 2016. Bone Res. 2016. PMID: 27563484 Free PMC article. Review.
-
MiR-26a Rescues Bone Regeneration Deficiency of Mesenchymal Stem Cells Derived From Osteoporotic Mice.Mol Ther. 2015 Aug;23(8):1349-1357. doi: 10.1038/mt.2015.101. Epub 2015 Jun 8. Mol Ther. 2015. PMID: 26050992 Free PMC article.
-
Modulation of Bone Morphogenetic Protein (BMP) 2 gene expression by Sp1 transcription factors.Gene. 2007 May 1;392(1-2):221-9. doi: 10.1016/j.gene.2006.12.032. Epub 2007 Jan 20. Gene. 2007. PMID: 17317039 Free PMC article.
-
MicroRNA-322 (miR-322) and its target protein Tob2 modulate Osterix (Osx) mRNA stability.J Biol Chem. 2013 May 17;288(20):14264-14275. doi: 10.1074/jbc.M112.432104. Epub 2013 Apr 5. J Biol Chem. 2013. PMID: 23564456 Free PMC article.
-
The estrogen receptor-alpha in osteoclasts mediates the protective effects of estrogens on cancellous but not cortical bone.Mol Endocrinol. 2010 Feb;24(2):323-34. doi: 10.1210/me.2009-0354. Epub 2010 Jan 6. Mol Endocrinol. 2010. PMID: 20053716 Free PMC article.
References
-
- Manolagas, S. C. (2000) Endocr. Rev. 21, 115-137. - PubMed
-
- NIH Consensus Development Panel on Osteoporosis Prevention, Diagnosis and Therapy (2001) J. Am. Med. Assoc. 285, 785-795. - PubMed
-
- Rodan, G. A. & Martin, T. J. (2000) Science 289, 1508-1514. - PubMed
-
- Karsenty, G. (1999) Genes Dev. 13, 3037-3051. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
