

Role of the yeast Rrp1 protein in the dynamics of pre-ribosome maturation
- PMID: 15100437
- PMCID: PMC1370572
- DOI: 10.1261/rna.5255804
Role of the yeast Rrp1 protein in the dynamics of pre-ribosome maturation
Abstract
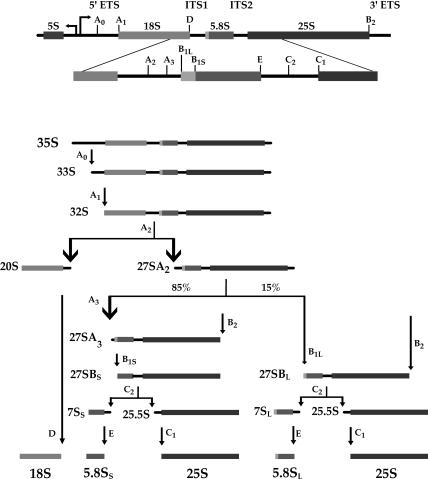
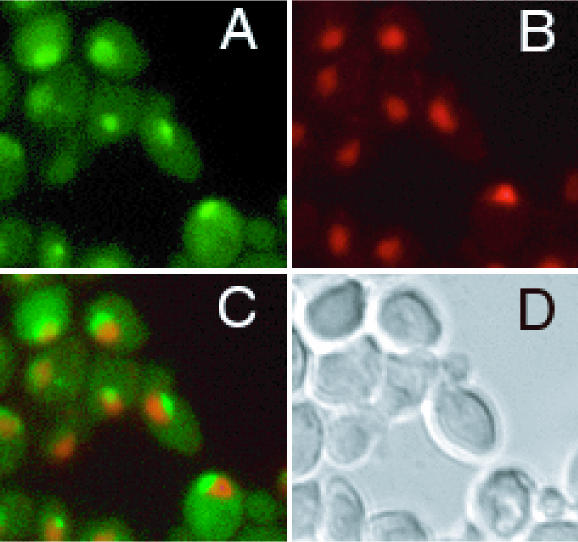
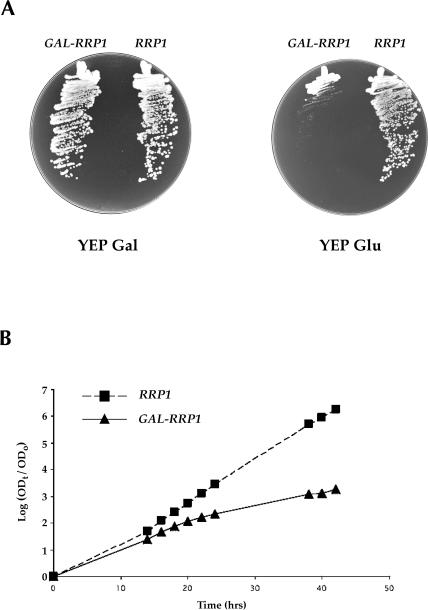
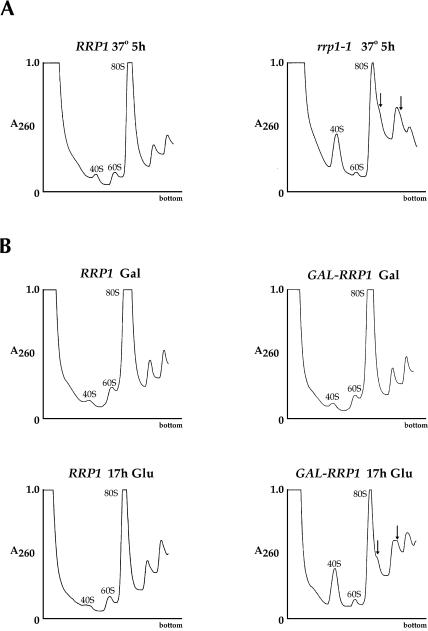
The Saccharomyces cerevisiae gene RRP1 encodes an essential, evolutionarily conserved protein necessary for biogenesis of 60S ribosomal subunits. Processing of 27S pre-ribosomal RNA to mature 25S rRNA is blocked and 60S subunits are deficient in the temperature-sensitive rrp1-1 mutant. We have used recent advances in proteomic analysis to examine in more detail the function of Rrp1p in ribosome biogenesis. We show that Rrp1p is a nucleolar protein associated with several distinct 66S pre-ribosomal particles. These pre-ribosomes contain ribosomal proteins plus at least 28 nonribosomal proteins necessary for production of 60S ribosomal subunits. Inactivation of Rrp1p inhibits processing of 27SA(3) to 27SB(S) pre-rRNA and of 27SB pre-rRNA to 7S plus 25.5S pre-rRNA. Thus, in the rrp1-1 mutant, 66S pre-ribosomal particles accumulate that contain 27SA(3) and 27SB(L) pre-ribosomal RNAs.
Figures

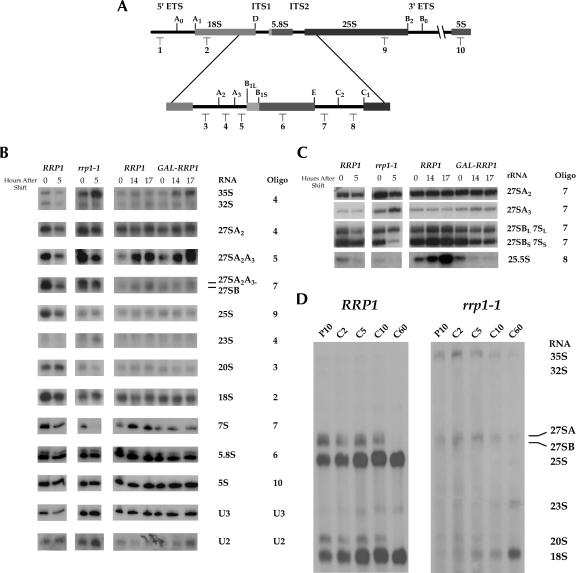

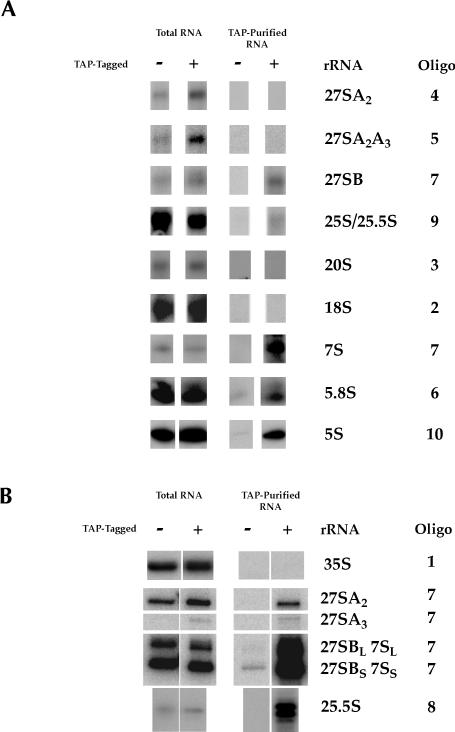
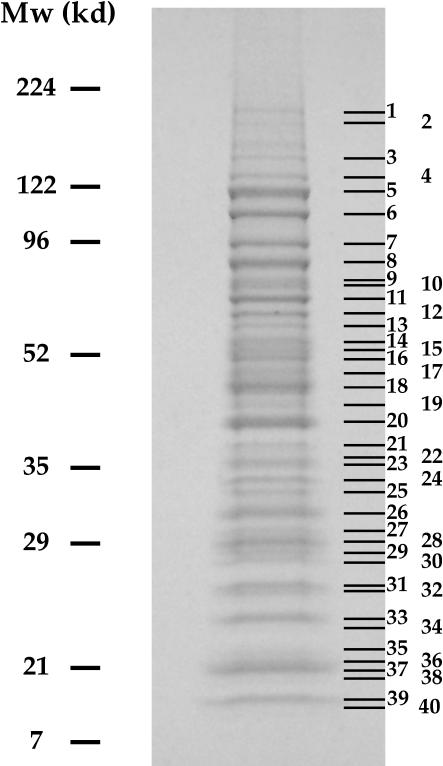
References
-
- Andersen, J.S., Lyon, C.E., Fox, A.H., Leung, A.K.L., Lam, Y.W., Steen, H., Mann, M., and Lamond, A.I. 2002. Directed proteomic analysis of the human nucleolus. Curr. Biol. 12: 1–11. - PubMed
-
- Andrew, C., Hopper, A.K., and Hall, B.D. 1976. A yeast mutant defective in the processing of 27S rRNA precursor. Mol. Gen. Genet. 144: 29–37. - PubMed
-
- Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., and Struhl, K. 1994. Current protocols in molecular biology. Wiley, New York.
-
- Baßler, J., Grandi, P., Gadal, O., Lessmann, T., Petfalski, E., Tollervey, D., Lechner, J., and Hurt, E. 2001. Identification of a 60S pre-ribosomal particle that is closely linked to nuclear export. Mol. Cell 8: 517–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases