

Comparative analysis detects dependencies among the 5' splice-site positions
- PMID: 15100438
- PMCID: PMC1370573
- DOI: 10.1261/rna.5196404
Comparative analysis detects dependencies among the 5' splice-site positions
Abstract
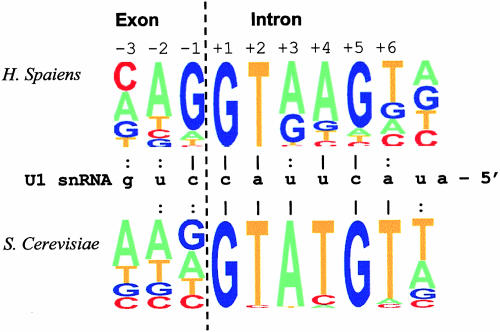
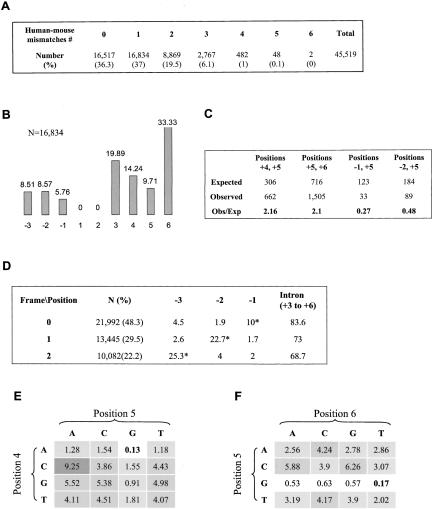
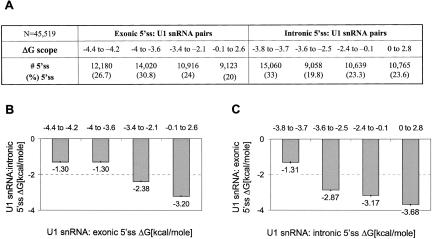
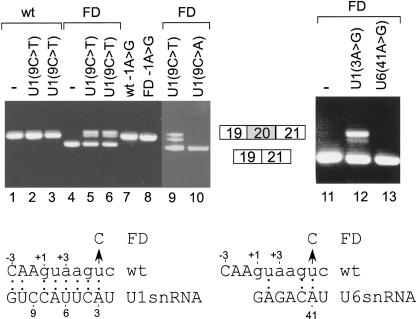
Human-mouse comparative genomics is an informative tool to assess sequence functionality as inferred from its conservation level. We used this approach to examine dependency among different positions of the 5' splice site. We compiled a data set of 50,493 homologous human-mouse internal exons and analyzed the frequency of changes among different positions of homologous human-mouse 5' splice-site pairs. We found mutual relationships between positions +4 and +5, +5 and +6, -2 and +5, and -1 and +5. We also demonstrated the association between the exonic and the intronic positions of the 5' splice site, in which a stronger interaction of U1 snRNA and the intronic portion of the 5' splice site compensates for weak interaction of U1 snRNA and the exonic portion of the 5' splice site, and vice versa. By using an ex vivo system that mimics the effect of mutation in the 5' splice site leading to familial dysautonomia, we demonstrated that U1 snRNA base-pairing with positions +6 and -1 is the only functional requirement for mRNA splicing of this 5' splice site. Our findings indicate the importance of U1 snRNA base-pairing to the exonic portion of the 5' splice site.
Figures

Similar articles
-
3' splice site recognition in S. cerevisiae does not require base pairing with U1 snRNA.Cell. 1993 May 21;73(4):803-12. doi: 10.1016/0092-8674(93)90258-r. Cell. 1993. PMID: 8500172
-
Exon mutations uncouple 5' splice site selection from U1 snRNA pairing.Cell. 1990 Nov 2;63(3):619-29. doi: 10.1016/0092-8674(90)90457-p. Cell. 1990. PMID: 2225068
-
Who's on first? The U1 snRNP-5' splice site interaction and splicing.Trends Biochem Sci. 1991 May;16(5):187-90. doi: 10.1016/0968-0004(91)90073-5. Trends Biochem Sci. 1991. PMID: 1882420 Review.
-
Intronic PAH gene mutations cause a splicing defect by a novel mechanism involving U1snRNP binding downstream of the 5' splice site.PLoS Genet. 2018 Apr 23;14(4):e1007360. doi: 10.1371/journal.pgen.1007360. eCollection 2018 Apr. PLoS Genet. 2018. PMID: 29684050 Free PMC article.
-
Can donor splice site recognition occur without the involvement of U1 snRNP?Biochem Soc Trans. 2008 Jun;36(Pt 3):548-50. doi: 10.1042/BST0360548. Biochem Soc Trans. 2008. PMID: 18482005 Review.
Cited by
-
Novel immunomodulators from hard ticks selectively reprogramme human dendritic cell responses.PLoS Pathog. 2013;9(6):e1003450. doi: 10.1371/journal.ppat.1003450. Epub 2013 Jun 27. PLoS Pathog. 2013. PMID: 23825947 Free PMC article.
-
A comprehensive characterization of cis-acting splicing-associated variants in human cancer.Genome Res. 2018 Aug;28(8):1111-1125. doi: 10.1101/gr.231951.117. Epub 2018 Jul 16. Genome Res. 2018. PMID: 30012835 Free PMC article.
-
Phylogenetically widespread alternative splicing at unusual GYNGYN donors.Genome Biol. 2006;7(7):R65. doi: 10.1186/gb-2006-7-7-R65. Genome Biol. 2006. PMID: 16869967 Free PMC article.
-
The "alternative" choice of constitutive exons throughout evolution.PLoS Genet. 2007 Nov;3(11):e203. doi: 10.1371/journal.pgen.0030203. PLoS Genet. 2007. PMID: 18020709 Free PMC article.
-
Improved identification of conserved cassette exons using Bayesian networks.BMC Bioinformatics. 2008 Nov 12;9:477. doi: 10.1186/1471-2105-9-477. BMC Bioinformatics. 2008. PMID: 19014490 Free PMC article.
References
-
- Aebi, M., Hornig, H., and Weissmann, C. 1987. 5′ cleavage site in eukaryotic pre-mRNA splicing is determined by the overall 5′ splice region, not by the conserved 5′ GU. Cell 50: 237–246. - PubMed
-
- Barrass, J.D. and Beggs, J.D. 2003. Splicing goes global. Trends Genet. 19: 295–298. - PubMed
-
- Brow, D.A. 2002. Allosteric cascade of spliceosome activation. Annu. Rev. Genet. 36: 333–360. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources