

Differential regulation of beta-defensin gene expression during Cryptosporidium parvum infection
- PMID: 15102787
- PMCID: PMC387892
- DOI: 10.1128/IAI.72.5.2772-2779.2004
Differential regulation of beta-defensin gene expression during Cryptosporidium parvum infection
Abstract
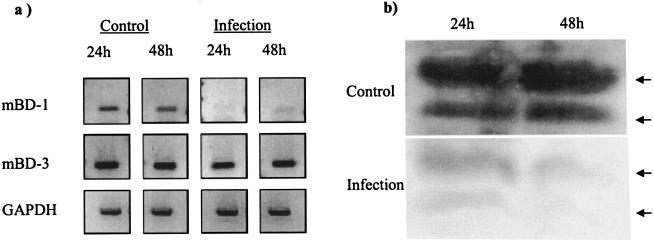
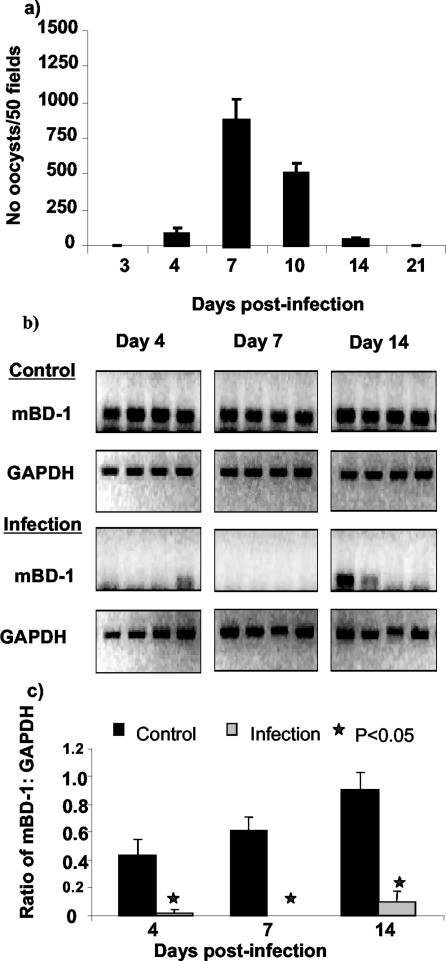
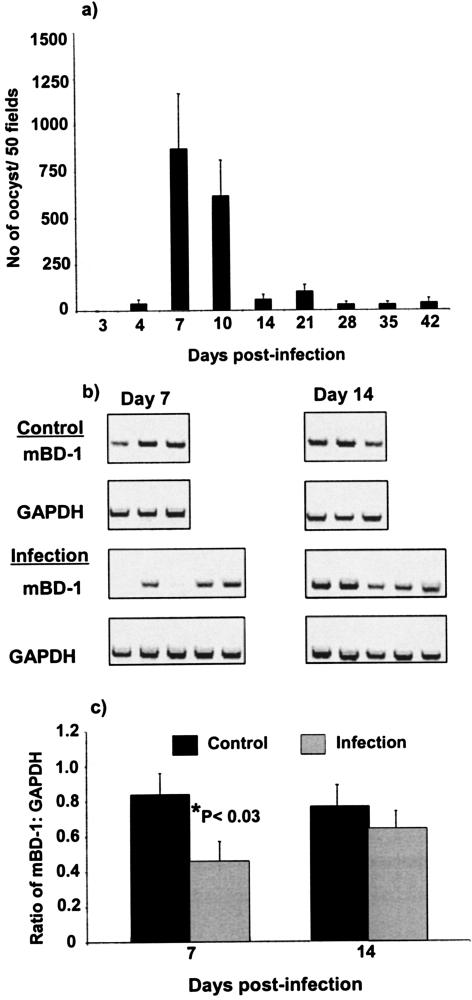
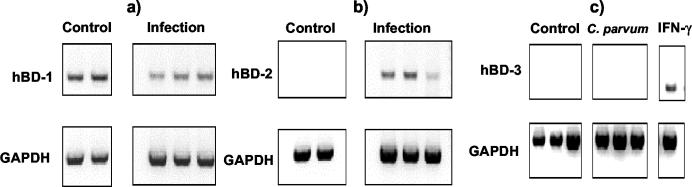
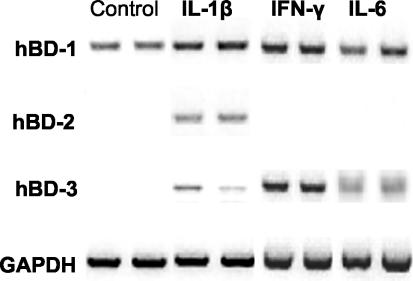
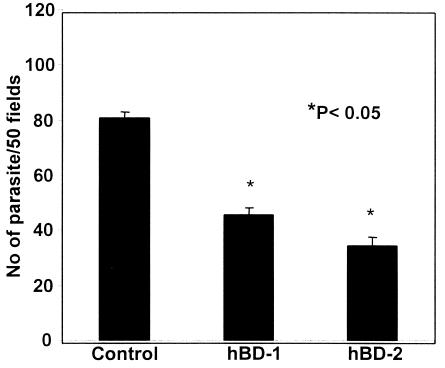
Invasion of enterocytes by pathogenic microbes evokes both innate and adaptive immune responses, and microbial pathogens have developed strategies to overcome the initial host immune defense. beta-Defensins are potentially important endogenous antibiotic-like effectors of innate immunity expressed by intestinal epithelia. In this study, the interplay between the enteric protozoan parasite Cryptosporidium parvum and host epithelial beta-defensin expression was investigated. Using human and murine models of infection, we demonstrated that C. parvum infection differentially regulates beta-defensin gene expression. Downregulation of murine beta-defensin-1 mRNA and protein was observed in both in vitro and in vivo models of infection. Infection of the human colonic HT29 cell line with the parasite resulted in differential effects on various members of the defensin gene family. Partial reduction in human beta-defensin-1 (hBD-1), induction of hBD-2, and no effect on hBD-3 gene expression was observed. Recombinant hBD-1 and hBD-2 peptides exhibited significant antimicrobial activity against C. parvum sporozoites in vitro. These findings demonstrate that C. parvum infection of enterocytes may affect the expression of various defensins in different ways and suggest that the overall outcome of the effect of antimicrobial peptides on early survival of the parasite may be complex.
Figures
Similar articles
-
Trans-suppression of defense DEFB1 gene in intestinal epithelial cells following Cryptosporidium parvum infection is associated with host delivery of parasite Cdg7_FLc_1000 RNA.Parasitol Res. 2018 Mar;117(3):831-840. doi: 10.1007/s00436-018-5759-0. Epub 2018 Jan 26. Parasitol Res. 2018. PMID: 29374323 Free PMC article.
-
Enteric beta-defensin: molecular cloning and characterization of a gene with inducible intestinal epithelial cell expression associated with Cryptosporidium parvum infection.Infect Immun. 1998 Mar;66(3):1045-56. doi: 10.1128/IAI.66.3.1045-1056.1998. Infect Immun. 1998. PMID: 9488394 Free PMC article.
-
IL-18 cytokine levels modulate innate immune responses and cryptosporidiosis in mice.J Eukaryot Microbiol. 2015 Jan-Feb;62(1):44-50. doi: 10.1111/jeu.12164. Epub 2014 Oct 23. J Eukaryot Microbiol. 2015. PMID: 25155632
-
Innate immune responses against Cryptosporidium parvum infection.Parasite Immunol. 2013 Feb;35(2):55-64. doi: 10.1111/pim.12020. Parasite Immunol. 2013. PMID: 23173616 Review.
-
Pathogenesis of Cryptosporidium parvum infection.Microbes Infect. 1999 Feb;1(2):141-8. doi: 10.1016/s1286-4579(99)80005-7. Microbes Infect. 1999. PMID: 10594978 Review.
Cited by
-
The Mucosal Innate Immune Response to Cryptosporidium parvum, a Global One Health Issue.Front Cell Infect Microbiol. 2021 May 25;11:689401. doi: 10.3389/fcimb.2021.689401. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34113580 Free PMC article. Review.
-
Regulation of antimicrobial peptide gene expression by nutrients and by-products of microbial metabolism.Eur J Nutr. 2012 Dec;51(8):899-907. doi: 10.1007/s00394-012-0415-4. Epub 2012 Jul 15. Eur J Nutr. 2012. PMID: 22797470 Free PMC article. Review.
-
Recombinant human beta 2-defensin fusion proteins as a tool to investigate defensin structure and function in small human intestinal tissue samples.Inflamm Res. 2012 Dec;61(12):1411-20. doi: 10.1007/s00011-012-0544-5. Epub 2012 Aug 26. Inflamm Res. 2012. PMID: 22922953
-
Genome-wide immunity studies in the rabbit: transcriptome variations in peripheral blood mononuclear cells after in vitro stimulation by LPS or PMA-Ionomycin.BMC Genomics. 2015 Jan 23;16(1):26. doi: 10.1186/s12864-015-1218-9. BMC Genomics. 2015. PMID: 25613284 Free PMC article.
-
Hide-and-Seek: A Game Played between Parasitic Protists and Their Hosts.Microorganisms. 2021 Nov 25;9(12):2434. doi: 10.3390/microorganisms9122434. Microorganisms. 2021. PMID: 34946036 Free PMC article. Review.
References
-
- Ayabe, T., D. P. Satchell, C. L. Wilson, W. C. Parks, M. E. Selsted, and A. J. Ouellette. 2000. Secretion of microbicidal alpha-defensins by intestinal Paneth cells in response to bacteria. Nat. Immunol. 1:113-118. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
