

Mutations in Caenorhabditis elegans cytoplasmic dynein components reveal specificity of neuronal retrograde cargo
- PMID: 15102906
- PMCID: PMC6729415
- DOI: 10.1523/JNEUROSCI.5039-03.2004
Mutations in Caenorhabditis elegans cytoplasmic dynein components reveal specificity of neuronal retrograde cargo
Abstract
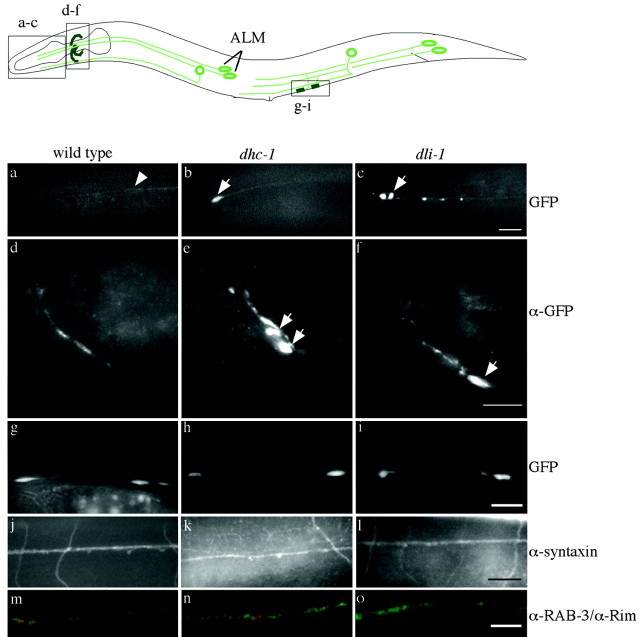
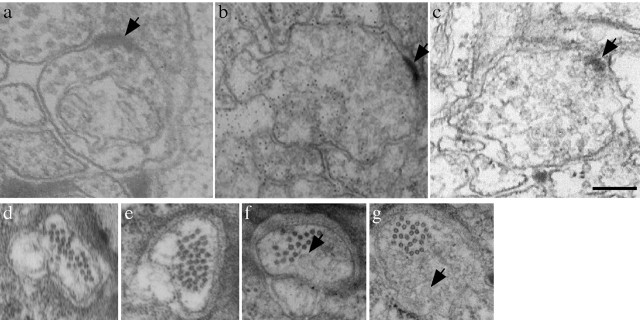
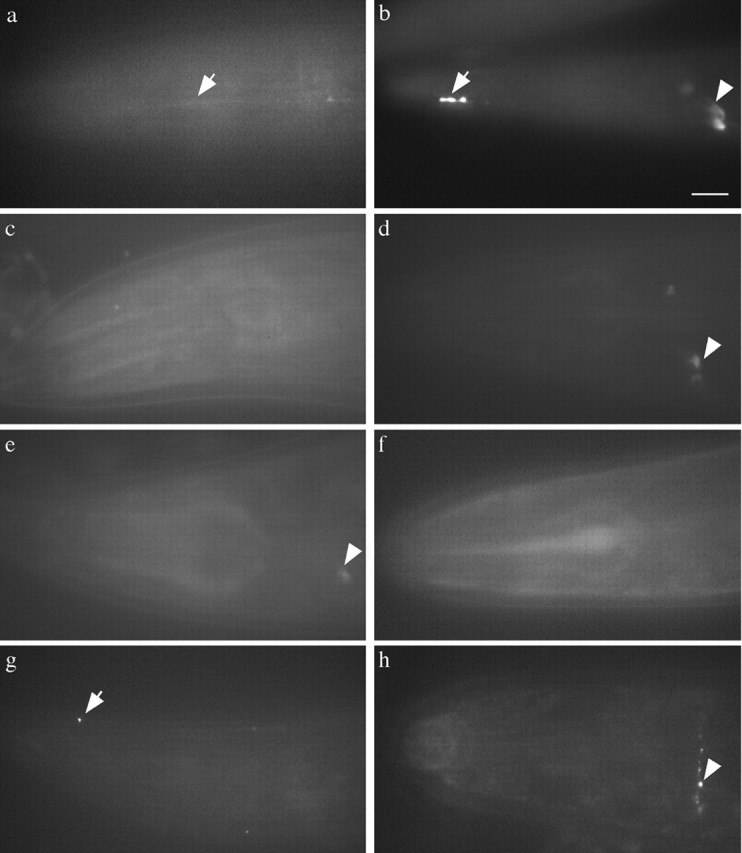
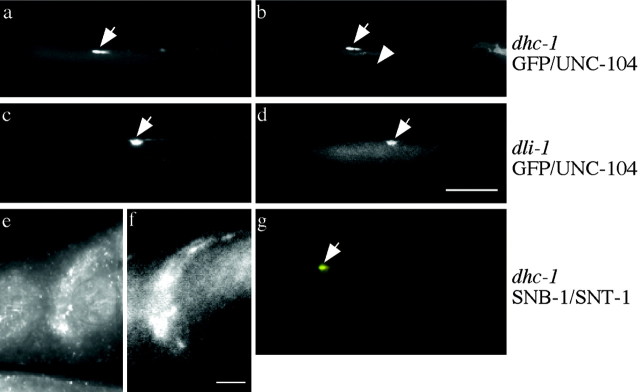
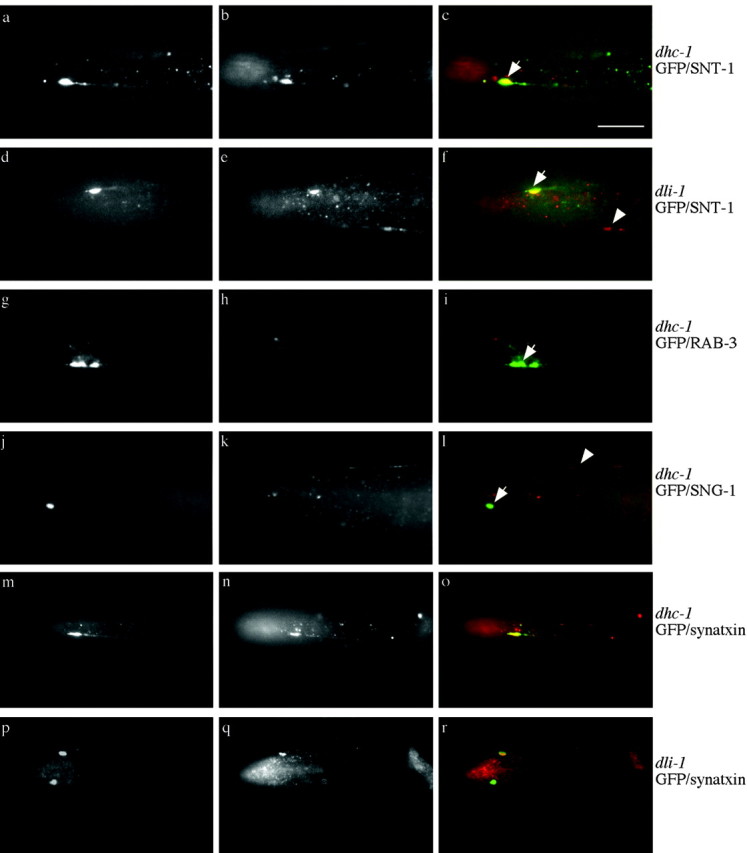
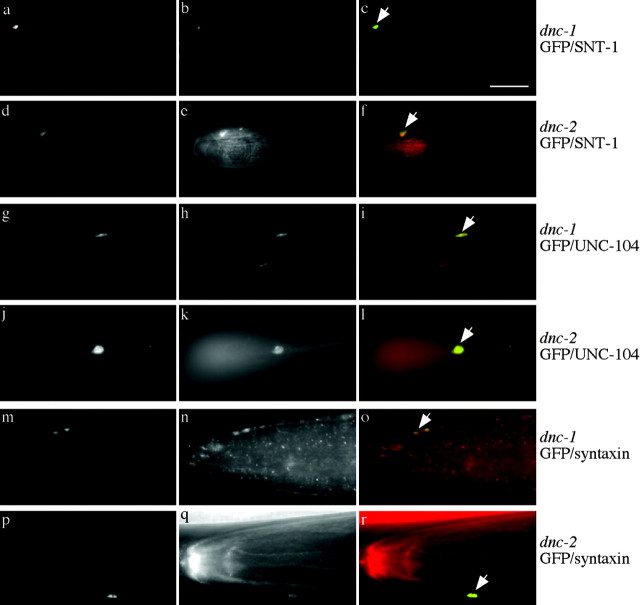
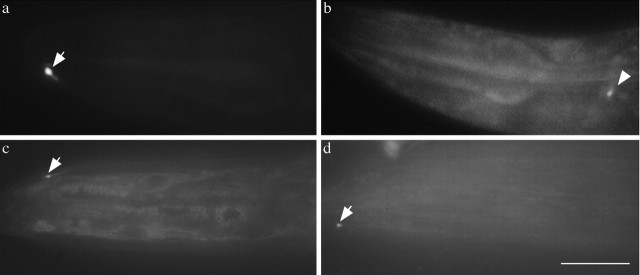
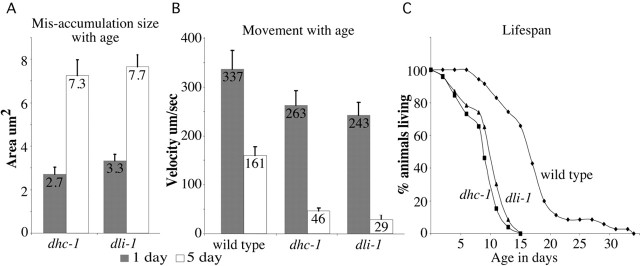
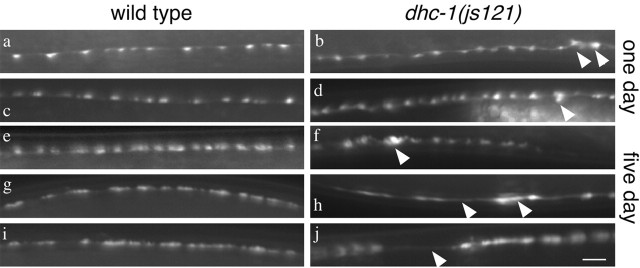
We describe Caenorhabditis elegans dynein complex mutants, which misaccumulate synaptic proteins at the ends of neuronal processes. Ultrastructural analysis revealed irregularly sized vesicles that likely represent accumulation of cargo. We propose that synaptobrevin, synaptotagmin, and UNC-104 are specific cargoes of the dynein complex. Many cargoes link to dynein via interactions between dynactin and vesicle-associated spectrin. However, loss of spectrin results in only mild and occasional defects in synaptobrevin localization. Thus, the dynein-dynactin complex shows neuronal cargo selectivity without spectrin being a critical component of cargo binding. We observed parallels to progressive motor neuron disease symptoms in these animals. With age, neuronal misaccumulations increase in size and frequency; locomotion becomes progressively slower; and life span is shortened. These mutants provide a model to assess whether defects in transport of specific cargo mediate neuronal dysfunction.
Figures
Similar articles
-
Insights on UNC-104-dynein/dynactin interactions and their implications on axonal transport in Caenorhabditis elegans.J Neurosci Res. 2019 Feb;97(2):185-201. doi: 10.1002/jnr.24339. Epub 2018 Oct 12. J Neurosci Res. 2019. PMID: 30311677
-
A transient helix in the disordered region of dynein light intermediate chain links the motor to structurally diverse adaptors for cargo transport.PLoS Biol. 2019 Jan 7;17(1):e3000100. doi: 10.1371/journal.pbio.3000100. eCollection 2019 Jan. PLoS Biol. 2019. PMID: 30615611 Free PMC article.
-
The lipid binding pleckstrin homology domain in UNC-104 kinesin is necessary for synaptic vesicle transport in Caenorhabditis elegans.Mol Biol Cell. 2004 Aug;15(8):3729-39. doi: 10.1091/mbc.e04-04-0326. Epub 2004 May 21. Mol Biol Cell. 2004. PMID: 15155810 Free PMC article.
-
Cytoplasmic dynein/dynactin function and dysfunction in motor neurons.Int J Dev Neurosci. 2006 Apr-May;24(2-3):103-11. doi: 10.1016/j.ijdevneu.2005.11.013. Epub 2006 Jan 6. Int J Dev Neurosci. 2006. PMID: 16406469 Review.
-
Controversies in synaptic vesicle exocytosis.J Cell Sci. 2003 Sep 15;116(Pt 18):3661-6. doi: 10.1242/jcs.00687. J Cell Sci. 2003. PMID: 12917353 Review. No abstract available.
Cited by
-
Fluorescence microscopy applied to intracellular transport by microtubule motors.J Biosci. 2018 Jul;43(3):437-445. J Biosci. 2018. PMID: 30002263 Review.
-
Differential regulation of polarized synaptic vesicle trafficking and synapse stability in neural circuit rewiring in Caenorhabditis elegans.PLoS Genet. 2017 Jun 21;13(6):e1006844. doi: 10.1371/journal.pgen.1006844. eCollection 2017 Jun. PLoS Genet. 2017. PMID: 28636662 Free PMC article.
-
Kinesin-3 and dynein cooperate in long-range retrograde endosome motility along a nonuniform microtubule array.Mol Biol Cell. 2011 Oct;22(19):3645-57. doi: 10.1091/mbc.E11-03-0217. Epub 2011 Aug 10. Mol Biol Cell. 2011. PMID: 21832152 Free PMC article.
-
UNC-16/JIP3 and UNC-76/FEZ1 limit the density of mitochondria in C. elegans neurons by maintaining the balance of anterograde and retrograde mitochondrial transport.Sci Rep. 2018 Jun 12;8(1):8938. doi: 10.1038/s41598-018-27211-9. Sci Rep. 2018. PMID: 29895958 Free PMC article.
-
Genetic analysis of a novel tubulin mutation that redirects synaptic vesicle targeting and causes neurite degeneration in C. elegans.PLoS Genet. 2014 Nov 13;10(11):e1004715. doi: 10.1371/journal.pgen.1004715. eCollection 2014 Nov. PLoS Genet. 2014. PMID: 25392990 Free PMC article.
References
-
- Bennett V, Baines AJ (2001) Spectrin and ankyrin-based pathways: metazoan inventions for integrating cells into tissues. Physiol Rev 81: 1353-1392. - PubMed
-
- Chalfie M (1993) Touch receptor development and function in Caenorhabditis elegans J Neurobiol 24: 1433-1441. - PubMed
-
- Clement AM, Nguyen MD, Roberts EA, Garcia ML, Boillee S, Rule M, Mc-Mahon AP, Doucette W, Siwek D, Ferrante RJ, Brown Jr RH, Julien JP, Goldstein LS, Cleveland DW (2003) Wild-type nonneuronal cells extend survival of SOD1 mutant motor neurons in ALS mice. Science 302: 113-117. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources