

Octasaccharide is the minimal length unit required for efficient binding of cyclophilin B to heparin and cell surface heparan sulphate
- PMID: 15109301
- PMCID: PMC1133832
- DOI: 10.1042/BJ20031453
Octasaccharide is the minimal length unit required for efficient binding of cyclophilin B to heparin and cell surface heparan sulphate
Abstract
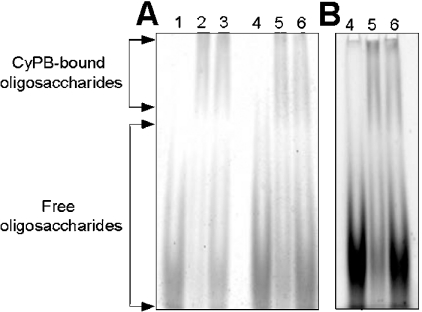
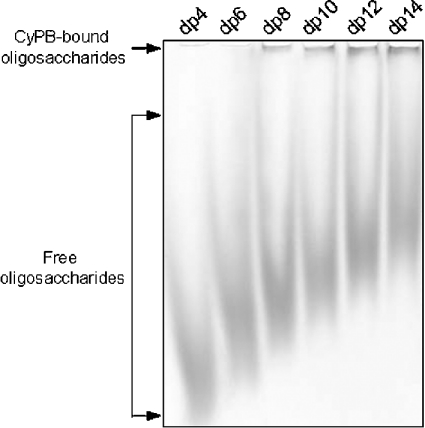
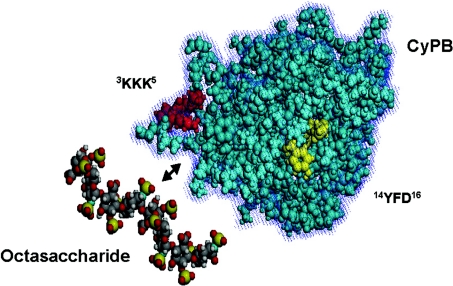
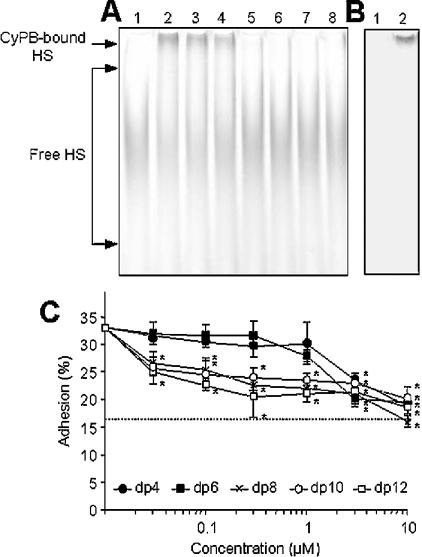
Cyclophilin B (CyPB) is a heparin-binding protein first identified as a receptor for cyclosporin A. In previous studies, we reported that CyPB triggers chemotaxis and integrin-mediated adhesion of T-lymphocytes by way of interaction with two types of binding sites. The first site corresponds to a signalling receptor; the second site has been identified as heparan sulphate (HS) and appears crucial to induce cell adhesion. Characterization of the HS-binding unit is critical to understand the requirement of HS in pro-adhesive activity of CyPB. By using a strategy based on gel mobility shift assays with fluorophore-labelled oligosaccharides, we demonstrated that the minimal heparin unit required for efficient binding of CyPB is an octasaccharide. The mutants CyPB(KKK-) [where KKK- refers to the substitutions K3A(Lys3-->Ala)/K4A/K5A] and CyPB(DeltaYFD) (where Tyr14-Phe-Asp16 has been deleted) failed to interact with octasaccharides, confirming that the Y14FD16 and K3KK5 clusters are required for CyPB binding. Molecular modelling revealed that both clusters are spatially arranged so that they may act synergistically to form a binding site for the octasaccharide. We then demonstrated that heparin-derived octasaccharides and higher degree of polymerization oligosaccharides inhibited the interaction between CyPB and fluorophore-labelled HS chains purified from T-lymphocytes, and strongly reduced the HS-dependent pro-adhesive activity of CyPB. However, oligosaccharides or heparin were unable to restore adhesion of heparinase-treated T-lymphocytes, indicating that HS has to be present on the cell membrane to support the pro-adhesive activity of CyPB. Altogether, these results demonstrate that the octasaccharide is likely to be the minimal length unit required for efficient binding of CyPB to cell surface HS and consequent HS-dependent cell responses.
Figures

Similar articles
-
Structural and functional characterization of the interaction between cyclophilin B and a heparin-derived oligosaccharide.J Biol Chem. 2007 Nov 23;282(47):34148-58. doi: 10.1074/jbc.M706353200. Epub 2007 Sep 12. J Biol Chem. 2007. PMID: 17855358
-
Interaction with glycosaminoglycans is required for cyclophilin B to trigger integrin-mediated adhesion of peripheral blood T lymphocytes to extracellular matrix.Proc Natl Acad Sci U S A. 2002 Mar 5;99(5):2714-9. doi: 10.1073/pnas.052284899. Epub 2002 Feb 26. Proc Natl Acad Sci U S A. 2002. PMID: 11867726 Free PMC article.
-
The heparin/heparan sulfate sequence that interacts with cyclophilin B contains a 3-O-sulfated N-unsubstituted glucosamine residue.J Biol Chem. 2007 Aug 17;282(33):24416-29. doi: 10.1074/jbc.M701835200. Epub 2007 Jun 22. J Biol Chem. 2007. PMID: 17588944
-
The role of heparin/heparan sulphate in the IFN-γ-led Arena.Biochimie. 2020 Mar;170:1-9. doi: 10.1016/j.biochi.2019.11.018. Epub 2019 Nov 30. Biochimie. 2020. PMID: 31794784 Review.
-
Using structurally defined oligosaccharides to understand the interactions between proteins and heparan sulfate.Curr Opin Struct Biol. 2018 Jun;50:155-161. doi: 10.1016/j.sbi.2018.04.003. Epub 2018 Apr 21. Curr Opin Struct Biol. 2018. PMID: 29684759 Free PMC article. Review.
Cited by
-
Participation of 3-O-sulfated heparan sulfates in the protection of macrophages by herpes simplex virus-1 glycoprotein D and cyclophilin B against apoptosis.FEBS Open Bio. 2016 Dec 24;7(2):133-148. doi: 10.1002/2211-5463.12145. eCollection 2017 Feb. FEBS Open Bio. 2016. PMID: 28174681 Free PMC article.
-
Extracellular cyclophilin-A stimulates ERK1/2 phosphorylation in a cell-dependent manner but broadly stimulates nuclear factor kappa B.Cancer Cell Int. 2012 Jul 4;12(1):19. doi: 10.1186/1475-2867-12-19. Cancer Cell Int. 2012. PMID: 22631225 Free PMC article.
-
Synthesis of heparan sulfate with cyclophilin B-binding properties is determined by cell type-specific expression of sulfotransferases.J Biol Chem. 2010 Jan 15;285(3):1701-15. doi: 10.1074/jbc.M109.018184. Epub 2009 Nov 23. J Biol Chem. 2010. PMID: 19940140 Free PMC article.
-
Preferential chemotaxis of activated human CD4+ T cells by extracellular cyclophilin A.J Leukoc Biol. 2007 Sep;82(3):613-8. doi: 10.1189/jlb.0506317. Epub 2007 May 31. J Leukoc Biol. 2007. PMID: 17540735 Free PMC article.
-
Differential contribution of the repeats to heparin binding of HBHA, a major adhesin of Mycobacterium tuberculosis.PLoS One. 2012;7(3):e32421. doi: 10.1371/journal.pone.0032421. Epub 2012 Mar 5. PLoS One. 2012. PMID: 22403657 Free PMC article.
References
-
- Handschumacher R. E., Harding M. W., Rice J., Drugge R. J., Speicher D. W. Cyclophilin: a specific cytosolic binding protein for cyclosporin A. Science. 1984;226:544–547. - PubMed
-
- Schreiber S. L. Chemistry and biology of the immunophilins and their immunosuppressive ligands. Science. 1991;251:283–287. - PubMed
-
- Fischer G., Wittmann-Liebold B., Lang K., Kiefhaber T., Schmid F. X. Cyclophilin and peptidyl-prolyl cis-trans isomerase are probably identical proteins. Nature (London) 1989;337:476–478. - PubMed
-
- Takahashi N., Hayano T., Suzuki M. Peptidyl-prolyl cis-trans isomerase is the cyclosporin A-binding protein cyclophilin. Nature (London) 1989;337:473–475. - PubMed
-
- Galat A. Variations of sequences and amino acid compositions of proteins that sustain their biological functions: an analysis of the cyclophilin family of proteins. Arch. Biochem. Biophys. 1999;371:149–162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical