

The roles of cis-inactivation by Notch ligands and of neuralized during eye and bristle patterning in Drosophila
- PMID: 15113404
- PMCID: PMC420236
- DOI: 10.1186/1471-213X-4-5
The roles of cis-inactivation by Notch ligands and of neuralized during eye and bristle patterning in Drosophila
Abstract
Background: The receptor protein Notch and its ligand Delta are expressed throughout proneural regions yet non-neural precursor cells are defined by Notch activity and neural precursor cells by Notch inactivity. Not even Delta overexpression activates Notch in neural precursor cells. It is possible that future neural cells are protected by cis-inactivation, in which ligands block activation of Notch within the same cell. The Delta-ubiquitin ligase Neuralized has been proposed to antagonize cis-inactivation, favoring Notch activation. Cis-inactivation and role of Neuralized have not yet been studied in tissues where neural precursor cells are resistant to nearby Delta, however, such as the R8 cells of the eye or the bristle precursor cells of the epidermis.
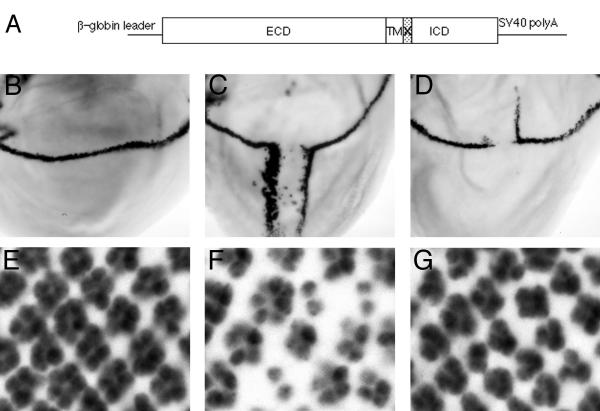
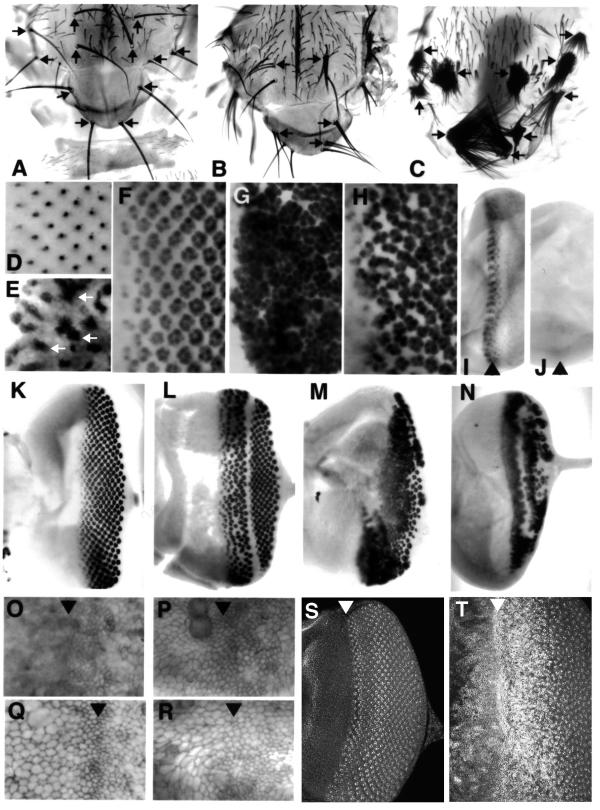
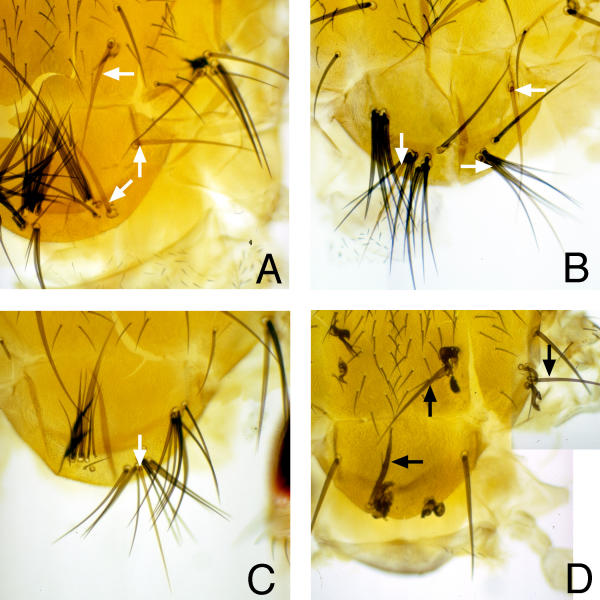
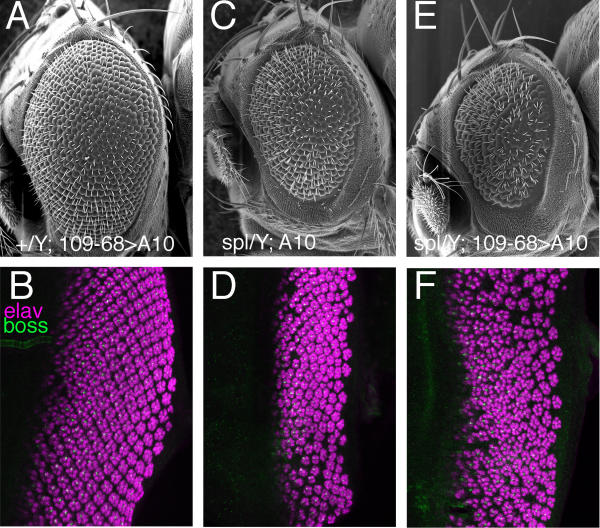
Results: Overexpressed ligands could block Notch signal transduction cell-autonomously in non-neural cells of the epidermis and retina, but did not activate Notch nonautonomously in neural cells. High ligand expression levels were required for cis-inactivation, and Serrate was more effective than Delta, although Delta is the ligand normally regulating neural specification. Differences between Serrate and Delta depended on the extracellular domains of the respective proteins. Neuralized was found to act cell nonautonomously in signal-sending cells during eye development, inconsistent with the view that Neuralized antagonizes cis-inactivation in non-neural cells.
Conclusions: Delta and Neuralized contribute cell nonautonomously to Notch signaling in neurogenesis, and the model that Neuralized antagonizes cis-inactivation to permit Notch activity and specification of non-neural cells is refuted. The molecular mechanism rendering Notch insensitive to paracrine activation in neural precursor cells remains uncertain.
Figures
Similar articles
-
The ubiquitin ligase Drosophila Mind bomb promotes Notch signaling by regulating the localization and activity of Serrate and Delta.Development. 2005 May;132(10):2319-32. doi: 10.1242/dev.01825. Epub 2005 Apr 13. Development. 2005. PMID: 15829515
-
The interplay between DSL proteins and ubiquitin ligases in Notch signaling.Development. 2005 Sep;132(18):4041-50. doi: 10.1242/dev.01979. Epub 2005 Aug 10. Development. 2005. PMID: 16093323
-
Distinct roles for Mind bomb, Neuralized and Epsin in mediating DSL endocytosis and signaling in Drosophila.Development. 2005 Jun;132(12):2883-94. doi: 10.1242/dev.01860. Development. 2005. PMID: 15930117
-
Delta-notch signaling and Drosophila cell fate choice.Dev Biol. 1994 Dec;166(2):415-30. doi: 10.1006/dbio.1994.1326. Dev Biol. 1994. PMID: 7813766 Review.
-
Multiple levels of Notch signal regulation (review).Mol Membr Biol. 2002 Jan-Mar;19(1):27-38. doi: 10.1080/09687680110112929. Mol Membr Biol. 2002. PMID: 11989820 Review.
Cited by
-
The Ubiquitin Conjugating Enzyme UbcD1 is Required for Notch Signaling Activation During Drosophila Wing Development.Front Genet. 2021 Oct 12;12:770853. doi: 10.3389/fgene.2021.770853. eCollection 2021. Front Genet. 2021. PMID: 34712275 Free PMC article.
-
Stochastic mechanisms of cell fate specification that yield random or robust outcomes.Annu Rev Cell Dev Biol. 2010;26:689-719. doi: 10.1146/annurev-cellbio-100109-104113. Annu Rev Cell Dev Biol. 2010. PMID: 20590453 Free PMC article. Review.
-
An extracellular region of Serrate is essential for ligand-induced cis-inhibition of Notch signaling.Development. 2013 May;140(9):2039-49. doi: 10.1242/dev.087916. Development. 2013. PMID: 23571220 Free PMC article.
-
Identification of genes that interact with Drosophila liquid facets.Genetics. 2007 Mar;175(3):1163-74. doi: 10.1534/genetics.106.067959. Epub 2006 Dec 18. Genetics. 2007. PMID: 17179082 Free PMC article.
-
Roles for the ubiquitin-proteasome pathway in protein quality control and signaling in the retina: implications in the pathogenesis of age-related macular degeneration.Mol Aspects Med. 2012 Aug;33(4):446-66. doi: 10.1016/j.mam.2012.04.001. Epub 2012 Apr 10. Mol Aspects Med. 2012. PMID: 22521794 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
