

Migration of antigen-specific T cells away from CXCR4-binding human immunodeficiency virus type 1 gp120
- PMID: 15113900
- PMCID: PMC400356
- DOI: 10.1128/jvi.78.10.5184-5193.2004
Migration of antigen-specific T cells away from CXCR4-binding human immunodeficiency virus type 1 gp120
Abstract
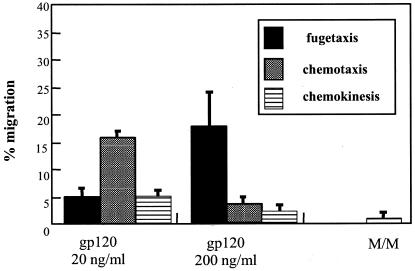
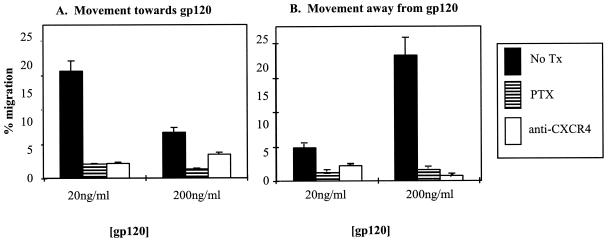
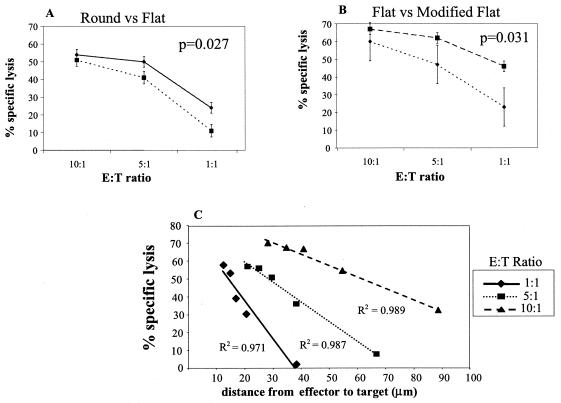
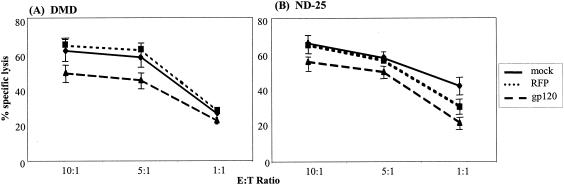
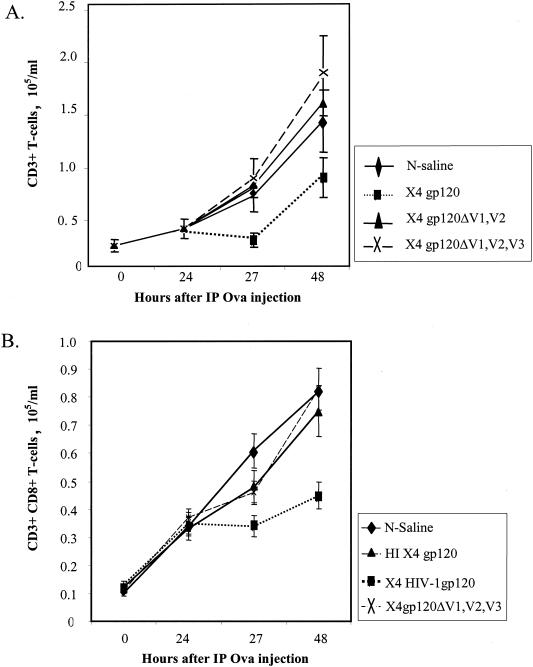
Cell-mediated immunity depends in part on appropriate migration and localization of cytotoxic T lymphocytes (CTL), a process regulated by chemokines and adhesion molecules. Many viruses, including human immunodeficiency virus type 1 (HIV-1), encode chemotactically active proteins, suggesting that dysregulation of immune cell trafficking may be a strategy for immune evasion. HIV-1 gp120, a retroviral envelope protein, has been shown to act as a T-cell chemoattractant via binding to the chemokine receptor and HIV-1 coreceptor CXCR4. We have previously shown that T cells move away from the chemokine stromal cell-derived factor 1 (SDF-1) in a concentration-dependent and CXCR4 receptor-mediated manner. Here, we demonstrate that CXCR4-binding HIV-1 X4 gp120 causes the movement of T cells, including HIV-specific CTL, away from high concentrations of the viral protein. This migratory response is CD4 independent and inhibited by anti-CXCR4 antibodies and pertussis toxin. Additionally, the expression of X4 gp120 by target cells reduces CTL efficacy in an in vitro system designed to account for the effect of cell migration on the ability of CTL to kill their target cells. Recombinant X4 gp120 also significantly reduced antigen-specific T-cell infiltration at a site of antigen challenge in vivo. The repellant activity of HIV-1 gp120 on immune cells in vitro and in vivo was shown to be dependent on the V2 and V3 loops of HIV-1 gp120. These data suggest that the active movement of T cells away from CXCR4-binding HIV-1 gp120, which we previously termed fugetaxis, may provide a novel mechanism by which HIV-1 evades challenge by immune effector cells in vivo.
Figures
References
-
- Addo, M. M., X. G. Yu, A. Rathod, D. Cohen, R. L. Eldridge, D. Strick, M. N. Johnston, C. Corcoran, A. G. Wurcel, C. A. Fitzpatrick, M. E. Feeney, W. R. Rodriguez, N. Basgoz, R. Draenert, D. R. Stone, C. Brander, P. J. Goulder, E. S. Rosenberg, M. Altfeld, and B. D. Walker. 2003. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 77:2081-2092. - PMC - PubMed
-
- Aliberti, J., J. G. Valenzuela, V. B. Carruthers, S. Hieny, J. Andersen, H. Charest, C. Reis e Sousa, A. Fairlamb, J. M. Ribeiro, and A. Sher. 2003. Molecular mimicry of a CCR5 binding-domain in the microbial activation of dendritic cells. Nat. Immunol. 4:485-490. - PubMed
-
- Alicami, A. 2003. Viral mimicry of cytokines, chemokines and their receptors. Nat. Rev. Immunol. 3:36-50. - PubMed
-
- Altfeld, M., J. van Lunzen, N. Frahm, X. G. Yu, C. Schneider, R. L. Eldridge, M. E. Feeney, D. Meyer-Olson, H.-J. Stellbrink, and B. D. Walker. 2002. Expansion of preexisting, lymph node-localized CD8+ T cells during supervised treatment interruptions in chronic HIV-1 infection. J. Clin. Investig. 109:837-843. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
