

Role of transmembrane domain and cytoplasmic tail amino acid sequences of influenza a virus neuraminidase in raft association and virus budding
- PMID: 15113907
- PMCID: PMC400379
- DOI: 10.1128/jvi.78.10.5258-5269.2004
Role of transmembrane domain and cytoplasmic tail amino acid sequences of influenza a virus neuraminidase in raft association and virus budding
Abstract
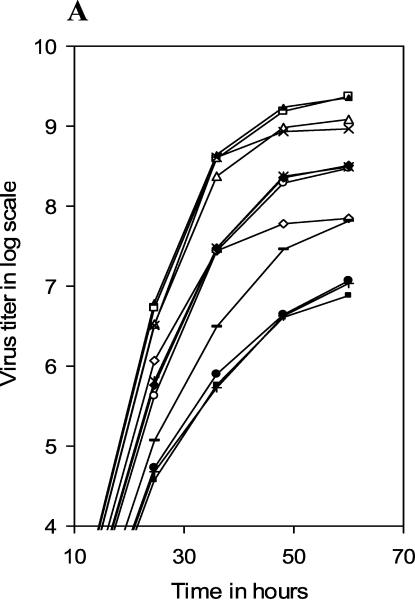
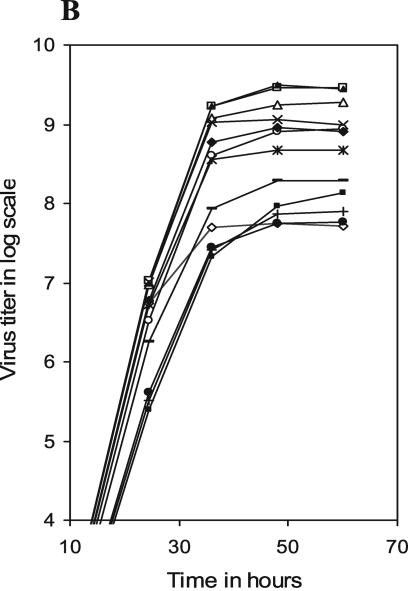
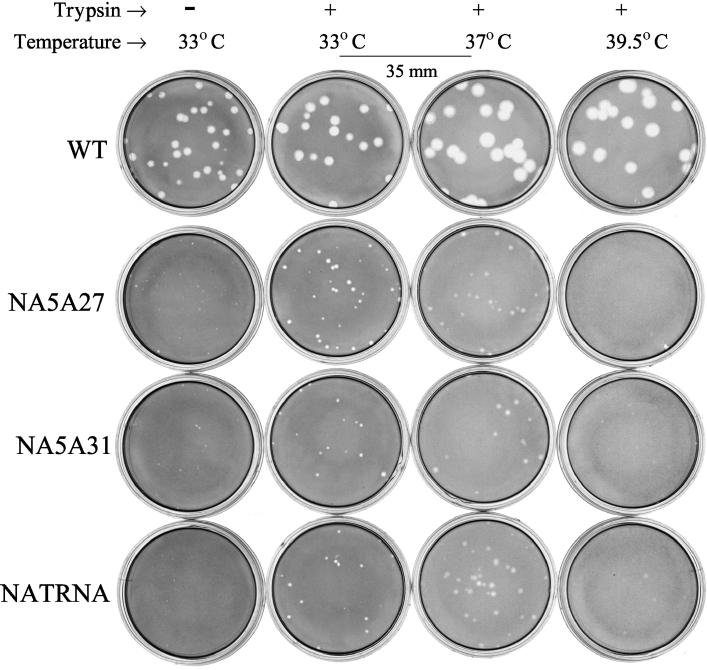
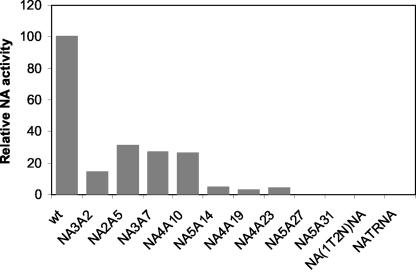
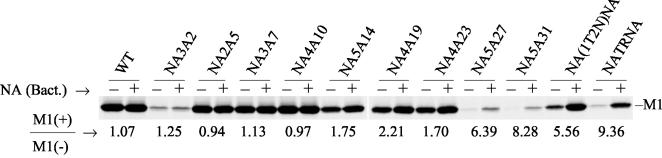
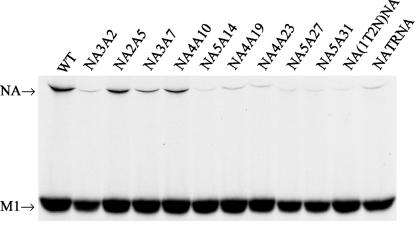
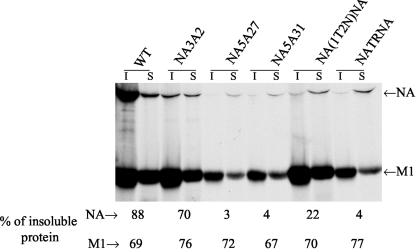
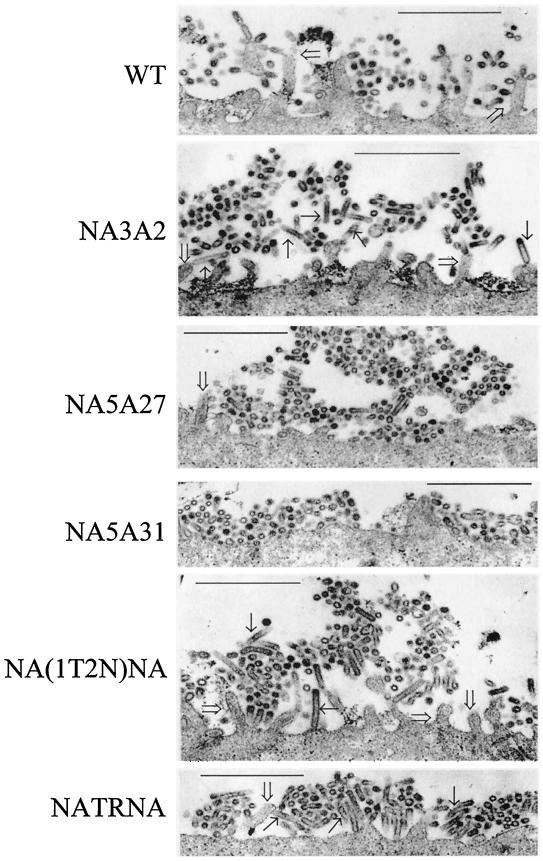
Influenza virus neuraminidase (NA), a type II transmembrane glycoprotein, possesses receptor-destroying activity and thereby facilitates virus release from the cell surface. Among the influenza A viruses, both the cytoplasmic tail (CT) and transmembrane domain (TMD) amino acid sequences of NA are highly conserved, yet their function(s) in virus biology remains unknown. To investigate the role of amino acid sequences of the CT and TMD on the virus life cycle, we systematically mutagenized the entire CT and TMD of NA by converting two to five contiguous amino acids to alanine. In addition, we also made two chimeric NA by replacing the CT proximal one-third amino acids of the NA TMD [NA(1T2N)NA] and the entire NA TMD (NATRNA) with that of human transferrin receptor (TR) (a type II transmembrane glycoprotein). We rescued transfectant mutant viruses by reverse genetics and examined their phenotypes. Our results show that all mutated and chimeric NAs could be rescued into transfectant viruses. Different mutants showed pleiotropic effects on virus growth and replication. Some mutants (NA2A5, NA3A7, and NA4A10) had little effect on virus growth while others (NA3A2, NA5A27, and NA5A31) produced about 50- to 100-fold-less infectious virus and still some others (NA5A14, NA4A19, and NA4A23) exhibited an intermediate phenotype. In general, mutations towards the ectodomain-proximal sequences of TMD progressively caused reduction in NA enzyme activity, affected lipid raft association, and attenuated virus growth. Electron microscopic analysis showed that these mutant viruses remained aggregated and bound to infected cell surfaces and could be released from the infected cells by bacterial NA treatment. Moreover, viruses containing mutations in the extreme N terminus of the CT (NA3A2) as well as chimeric NA containing the TMD replaced partially [NA(1T2N)NA] or fully (NATRNA) with TR TMD caused reduction in virus growth and exhibited the morphological phenotype of elongated particles. These results show that although the sequences of NA CT and TMD per se are not absolutely essential for the virus life cycle, specific amino acid sequences play a critical role in providing structural stability, enzyme activity, and lipid raft association of NA. In addition, aberrant morphogenesis including elongated particle formation of some mutant viruses indicates the involvement of NA in virus morphogenesis and budding.
Figures

References
-
- Air, G. M., and W. G. Laver. 1989. The neuraminidase of influenza virus. Proteins 6:341-356. - PubMed
-
- Barman, S., L. Adhikary, Y. Kawaoka, and D. P. Nayak. 2003. Influenza A virus hemagglutinin containing basolateral localization signal does not alter the apical budding of a recombinant influenza A virus in polarized MDCK cells. Virology 305:138-152. - PubMed
-
- Barman, S., A. Ali, E. K.-W. Hui, L. Adhikary, and D. P. Nayak. 2001. Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses. Virus Res. 77:61-69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
