

Deletion of Ser-171 causes inactivation, proteasome-mediated degradation and complete deficiency of human transaldolase
- PMID: 15115436
- PMCID: PMC1133831
- DOI: 10.1042/BJ20040413
Deletion of Ser-171 causes inactivation, proteasome-mediated degradation and complete deficiency of human transaldolase
Abstract
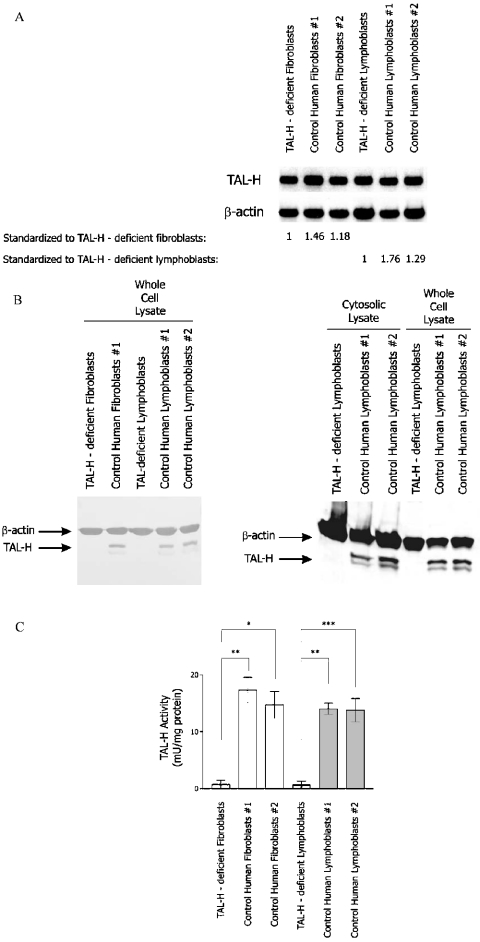
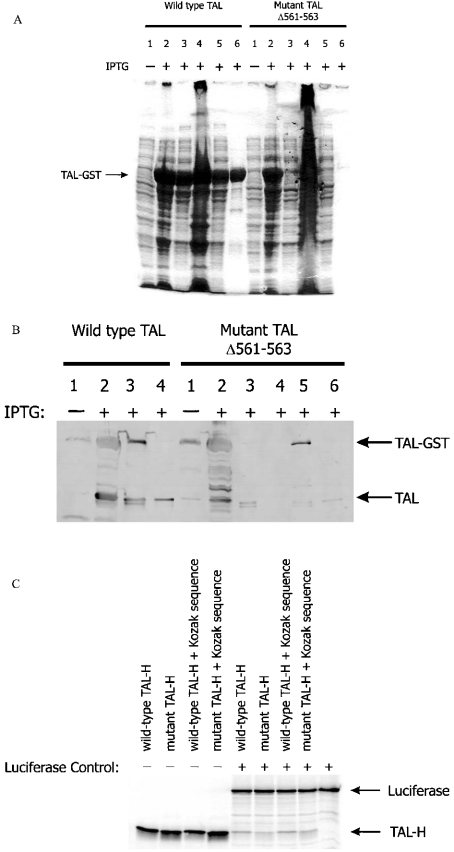
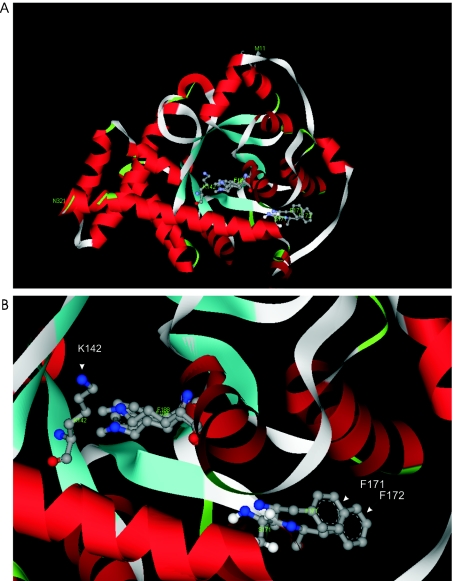
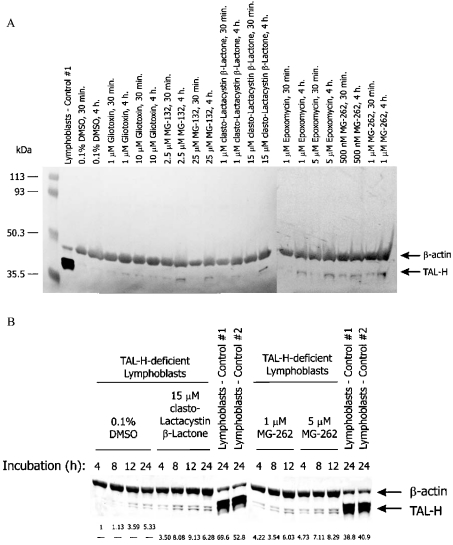
Homozygous deletion of three nucleotides coding for Ser-171 (S171) of TAL-H (human transaldolase) has been identified in a female patient with liver cirrhosis. Accumulation of sedoheptulose 7-phosphate raised the possibility of TAL (transaldolase) deficiency in this patient. In the present study, we show that the mutant TAL-H gene was effectively transcribed into mRNA, whereas no expression of the TALDeltaS171 protein or enzyme activity was detected in TALDeltaS171 fibroblasts or lymphoblasts. Unlike wild-type TAL-H-GST fusion protein (where GST stands for glutathione S-transferase), TALDeltaS171-GST was solubilized only in the presence of detergents, suggesting that deletion of Ser-171 caused conformational changes. Recombinant TALDeltaS171 had no enzymic activity. TALDeltaS171 was effectively translated in vitro using rabbit reticulocyte lysates, indicating that the absence of TAL-H protein in TALDeltaS171 fibroblasts and lymphoblasts may be attributed primarily to rapid degradation. Treatment with cell-permeable proteasome inhibitors led to the accumulation of TALDeltaS171 in whole cell lysates and cytosolic extracts of patient lymphoblasts, suggesting that deletion of Ser-171 led to rapid degradation by the proteasome. Although the TALDeltaS171 protein became readily detectable in proteasome inhibitor-treated cells, it displayed no appreciable enzymic activity. The results suggest that deletion of Ser-171 leads to inactivation and proteasome-mediated degradation of TAL-H. Since TAL-H is a regulator of apoptosis signal processing, complete deficiency of TAL-H may be relevant for the pathogenesis of liver cirrhosis.
Figures
References
-
- Mayes P. A. The pentose phosphate pathway and other pathways of hexose metabolism. In: Murray R. K., Granner D. K., Mayes P. A., Rodwell V. W., editors. Harper's Biochemistry. Norwalk, CT: Appleton and Lange; 1993. pp. 201–211.
-
- Cooper R. A., Bunn H. F. Hemolytic anemias. In: Wilson J. D., Braunwald E., Isselbacher K. J., Petersdorf R. G., Martin J. B., Fauci A. S., Root R. K., editors. Harrison's Principles of Internal Medicine. New York: McGraw-Hill; 1991. pp. 1531–1543.
-
- Banki K., Hutter E., Colombo E., Gonchoroff N. J., Perl A. Glutathione levels and sensitivity to apoptosis are regulated by changes in transaldolase expression. J. Biol. Chem. 1996;271:32994–33001. - PubMed
-
- Banki K., Hutter E., Gonchoroff N. J., Perl A. Molecular ordering in HIV-induced apoptosis: oxidative stress, activation of caspases, and cell survival are regulated by transaldolase. J. Biol. Chem. 1998;273:11944–11953. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous