

ClgR, a novel regulator of clp and lon expression in Streptomyces
- PMID: 15126487
- PMCID: PMC400634
- DOI: 10.1128/JB.186.10.3238-3248.2004
ClgR, a novel regulator of clp and lon expression in Streptomyces
Abstract
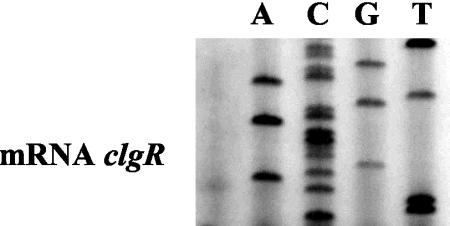
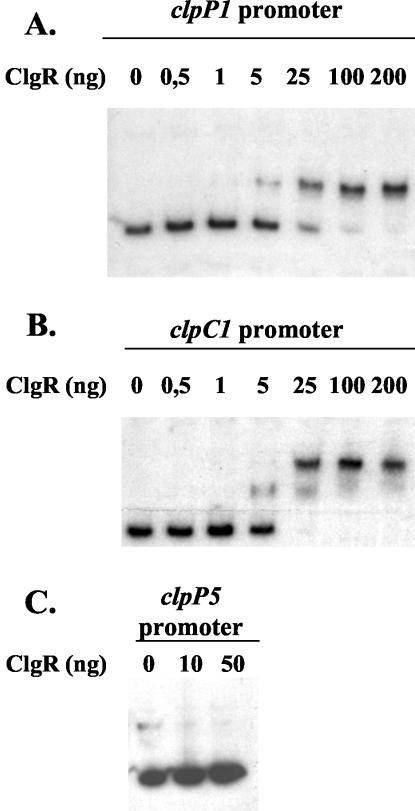
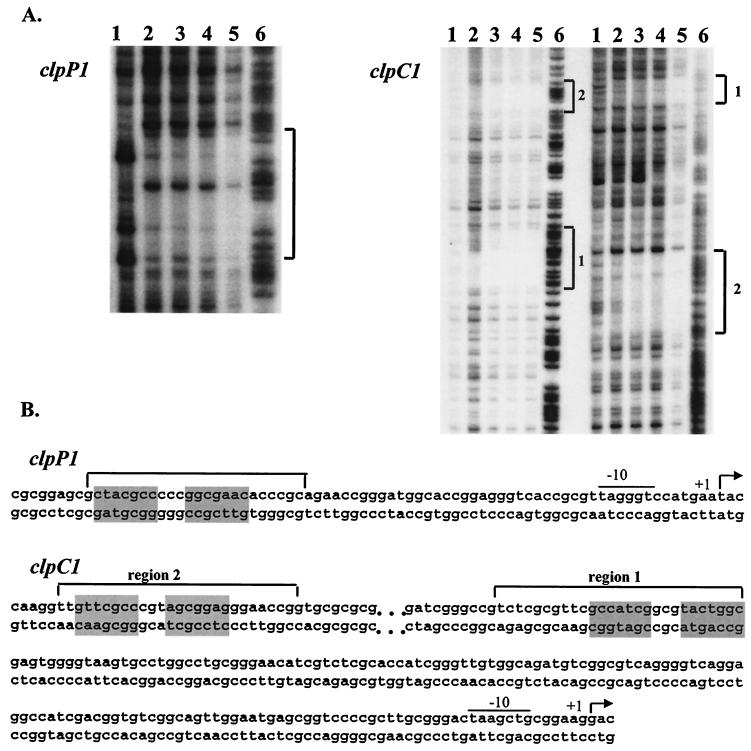
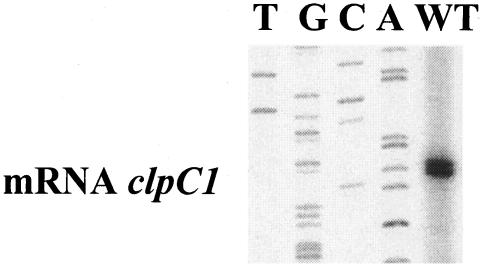
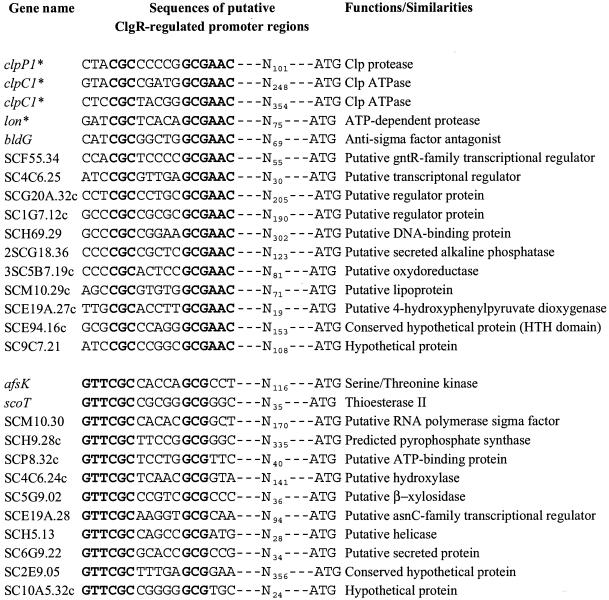
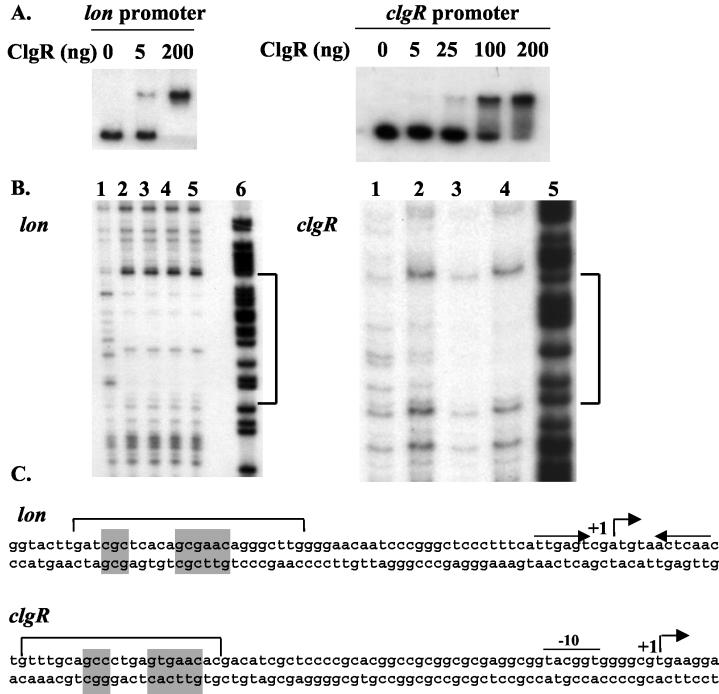
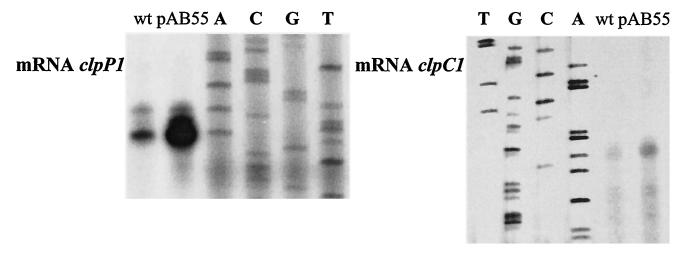
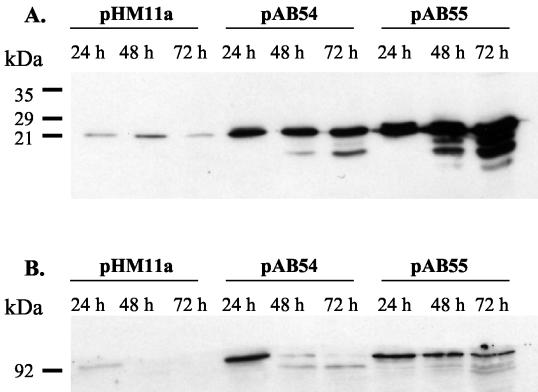
The clp genes encoding the Clp proteolytic complex are widespread among living organisms. Five clpP genes are present in Streptomyces. Among them, the clpP1 clpP2 operon has been shown to be involved in the Streptomyces growth cycle, as a mutation blocked differentiation at the substrate mycelium step. Four Clp ATPases have been identified in Streptomyces coelicolor (ClpX and three ClpC proteins) which are potential partners of ClpP1 ClpP2. The clpC1 gene appears to be essential, since no mutant has yet been obtained. clpP1 clpP2 and clpC1 are important for Streptomyces growth, and a study of their regulation is reported here. The clpP3 clpP4 operon, which has been studied in Streptomyces lividans, is induced in a clpP1 mutant strain, and regulation of its expression is mediated via PopR, a transcriptional regulator. We report here studies of clgR, a paralogue of popR, in S. lividans. Gel mobility shift assays and DNase I footprinting indicate that ClgR binds not only to the clpP1 and clpC1 promoters, but also to the promoter of the Lon ATP-dependent protease gene and the clgR promoter itself. ClgR recognizes the motif GTTCGC-5N-GCG. In vivo, ClgR acts as an activator of clpC1 gene and clpP1 operon expression. Similarly to PopR, ClgR degradation might be ClpP dependent and could be mediated via recognition of the two carboxy-terminal alanine residues.
Figures

References
-
- Amrein, K. E., B. Takacs, M. Stieger, J. Molnos, N. A. Flint, and P. Burn. 1995. Purification and characterization of recombinant human p50csk protein-tyrosine kinase from an Escherichia coli expression system overproducing the bacterial chaperones GroES and GroEL. Proc. Natl. Acad. Sci. USA 92:1048-1052. - PMC - PubMed
-
- Bentley, S. D., K. F. Chater, A. M. Cerdeno-Tarraga, G. L. Challis, N. R. Thomson, K. D. James, D. E. Harris, M. A. Quail, H. Kieser, D. Harper, A. Bateman, S. Brown, G. Chandra, C. W. Chen, M. Collins, A. Cronin, A. Fraser, A. Goble, J. Hidalgo, T. Hornsby, S. Howarth, C. H. Huang, T. Kieser, L. Larke, L. Murphy, K. Oliver, S. O'Neil, E. Rabbinowitsch, M. A. Rajandream, K. Rutherford, S. Rutter, K. Seeger, D. Saunders, S. Sharp, R. Squares, S. Squares, K. Taylor, T. Warren, A. Wietzorrek, J. Woodward, B. G. Barrell, J. Parkhill, and D. A. Hopwood. 2002. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 417:141-147. - PubMed
-
- Bradford, M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Chastanet, A., J. Fert, and T. Msadek. 2003. Comparative genomics reveal novel heat shock regulatory mechanisms in Staphylococcus aureus and other Gram-positive bacteria. Mol. Microbiol. 47:1061-1073. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
