

Two distinct pathways for trehalose assimilation in the yeast Saccharomyces cerevisiae
- PMID: 15128531
- PMCID: PMC404389
- DOI: 10.1128/AEM.70.5.2771-2778.2004
Two distinct pathways for trehalose assimilation in the yeast Saccharomyces cerevisiae
Abstract
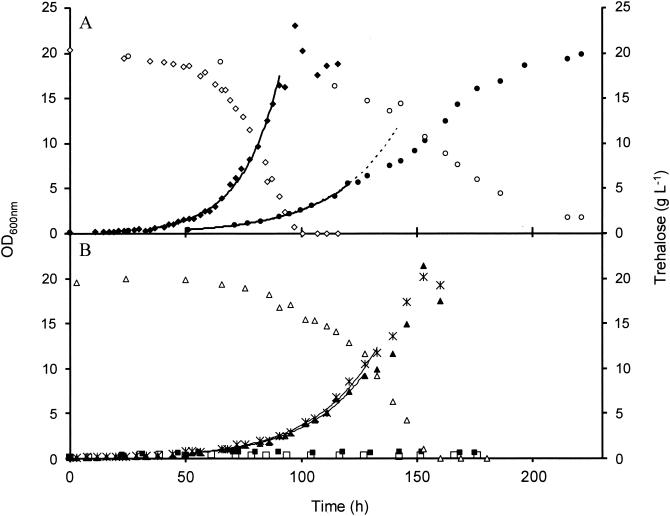
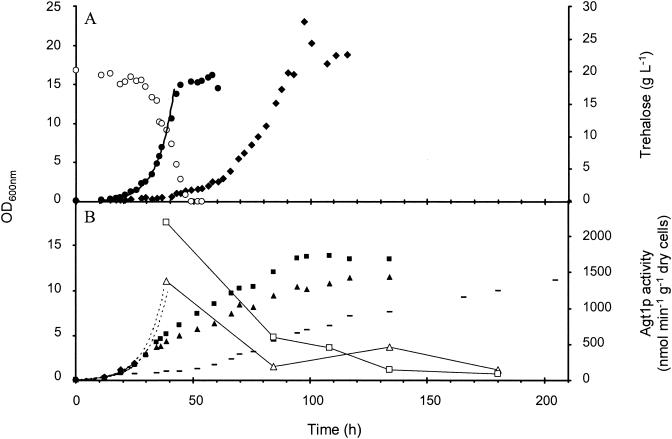
The yeast Saccharomyces cerevisiae can synthesize trehalose and also use this disaccharide as a carbon source for growth. However, the molecular mechanism by which extracellular trehalose can be transported to the vacuole and degraded by the acid trehalase Ath1p is not clear. By using an adaptation of the assay of invertase on whole cells with NaF, we showed that more than 90% of the activity of Ath1p is extracellular, splitting of the disaccharide into glucose. We also found that Agt1p-mediated trehalose transport and the hydrolysis of the disaccharide by the cytosolic neutral trehalase Nth1p are coupled and represent a second, independent pathway, although there are several constraints on this alternative route. First, the AGT1/MAL11 gene is controlled by the MAL system, and Agt1p was active in neither non-maltose-fermenting nor maltose-inducible strains. Second, Agt1p rapidly lost activity during growth on trehalose, by a mechanism similar to the sugar-induced inactivation of the maltose permease. Finally, both pathways are highly pH sensitive and effective growth on trehalose occurred only when the medium was buffered at around pH 5.0. The catabolism of trehalose was purely oxidative, and since levels of Ath1p limit the glucose flux in the cells, batch cultures on trehalose may provide a useful alternative to glucose-limited chemostat cultures for investigation of metabolic responses in yeast.
Figures
Similar articles
-
The three trehalases Nth1p, Nth2p and Ath1p participate in the mobilization of intracellular trehalose required for recovery from saline stress in Saccharomyces cerevisiae.Microbiology (Reading). 2009 Sep;155(Pt 9):3092-3099. doi: 10.1099/mic.0.024992-0. Epub 2009 Jun 11. Microbiology (Reading). 2009. PMID: 19520725
-
New insights into trehalose metabolism by Saccharomyces cerevisiae: NTH2 encodes a functional cytosolic trehalase, and deletion of TPS1 reveals Ath1p-dependent trehalose mobilization.Appl Environ Microbiol. 2008 Feb;74(3):605-14. doi: 10.1128/AEM.00557-07. Epub 2007 Dec 7. Appl Environ Microbiol. 2008. PMID: 18065618 Free PMC article.
-
The trehalose protective mechanism during thermal stress in Saccharomyces cerevisiae: the roles of Ath1 and Agt1.FEMS Yeast Res. 2018 Sep 1;18(6). doi: 10.1093/femsyr/foy066. FEMS Yeast Res. 2018. PMID: 30007297
-
Acid trehalase in yeasts and filamentous fungi: localization, regulation and physiological function.FEMS Yeast Res. 2005 Apr;5(6-7):503-11. doi: 10.1016/j.femsyr.2005.01.002. FEMS Yeast Res. 2005. PMID: 15780651 Review.
-
Molecular biology of trehalose and the trehalases in the yeast Saccharomyces cerevisiae.Prog Nucleic Acid Res Mol Biol. 1998;58:197-237. doi: 10.1016/s0079-6603(08)60037-9. Prog Nucleic Acid Res Mol Biol. 1998. PMID: 9308367 Review.
Cited by
-
Genomic content of a novel yeast species Hanseniaspora gamundiae sp. nov. from fungal stromata (Cyttaria) associated with a unique fermented beverage in Andean Patagonia, Argentina.PLoS One. 2019 Jan 30;14(1):e0210792. doi: 10.1371/journal.pone.0210792. eCollection 2019. PLoS One. 2019. PMID: 30699175 Free PMC article.
-
The secreted acid trehalase encoded by the CgATH1 gene is involved in Candida glabrata virulence.Mem Inst Oswaldo Cruz. 2020 Oct 30;115:e200401. doi: 10.1590/0074-02760200401. eCollection 2020. Mem Inst Oswaldo Cruz. 2020. PMID: 33146242 Free PMC article.
-
Kinetic Modeling of Saccharomyces cerevisiae Central Carbon Metabolism: Achievements, Limitations, and Opportunities.Metabolites. 2022 Jan 13;12(1):74. doi: 10.3390/metabo12010074. Metabolites. 2022. PMID: 35050196 Free PMC article. Review.
-
Improved Sugarcane-Based Fermentation Processes by an Industrial Fuel-Ethanol Yeast Strain.J Fungi (Basel). 2023 Jul 29;9(8):803. doi: 10.3390/jof9080803. J Fungi (Basel). 2023. PMID: 37623574 Free PMC article.
-
MAL62 overexpression and NTH1 deletion enhance the freezing tolerance and fermentation capacity of the baker's yeast in lean dough.Microb Cell Fact. 2016 Apr 4;15:54. doi: 10.1186/s12934-016-0453-3. Microb Cell Fact. 2016. PMID: 27039899 Free PMC article.
References
-
- Alizadeh, P., and D. J. Klionsky. 1996. Purification and biochemical characterization of the ATH1 gene product, vacuolar acid trehalase, from Saccharomyces cerevisiae. FEBS Lett. 391:273-278. - PubMed
-
- Attfield, P. V. 1997. Stress tolerance: the key to effective strains of industrial baker's yeast. Nat. Biotechnol. 15:1351-1357. - PubMed
-
- Barnett, J. A. 1997. Sugar utilization by Saccharomyces cerevisiae, p. 35-43. In F. K. Zimmermann and K. D. Entian (ed.), Yeast sugar metabolism. Technomic Publishing Co., Inc., Lancaster, Pa.
-
- Berben, G., J. Dumont, V. Gilliquet, P. A. Bolle, and F. Hilger. 1991. The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast 7:475-477. - PubMed
-
- Biswas, N., and A. K. Ghosh. 1998. Regulation of acid trehalase activity by association-dissociation in Saccharomyces cerevisiae. Biochim. Biophys. Acta 1379:245-256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
