

Glycoprotein 96 can chaperone both MHC class I- and class II-restricted epitopes for in vivo presentation, but selectively primes CD8+ T cell effector function
- PMID: 15128793
- PMCID: PMC2846363
- DOI: 10.4049/jimmunol.172.10.6087
Glycoprotein 96 can chaperone both MHC class I- and class II-restricted epitopes for in vivo presentation, but selectively primes CD8+ T cell effector function
Abstract
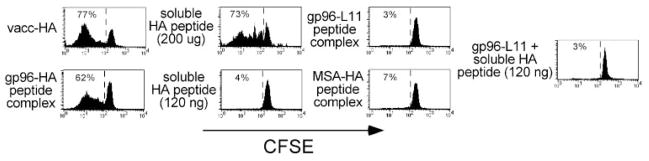
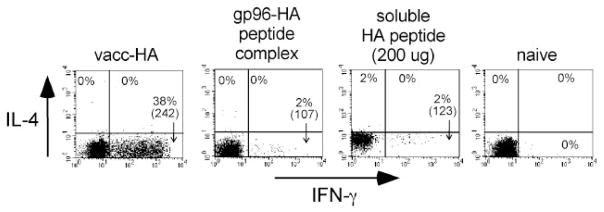
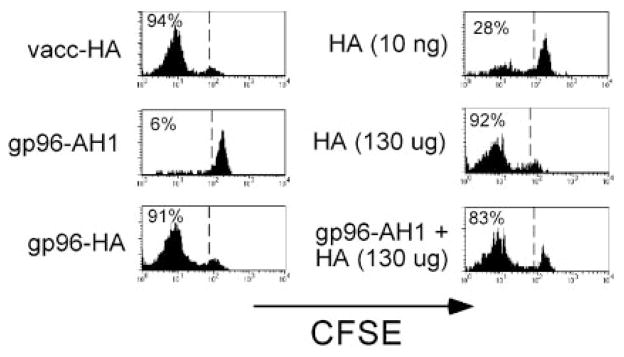
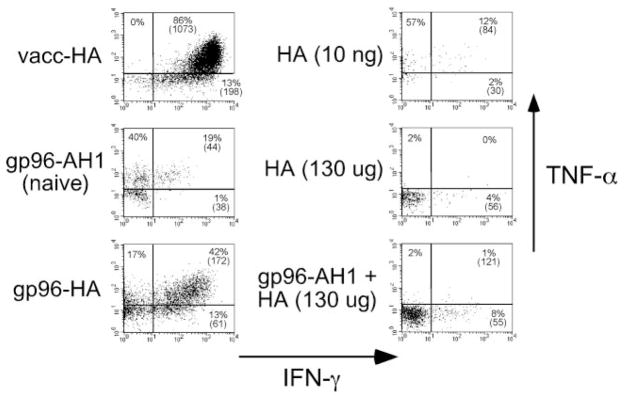
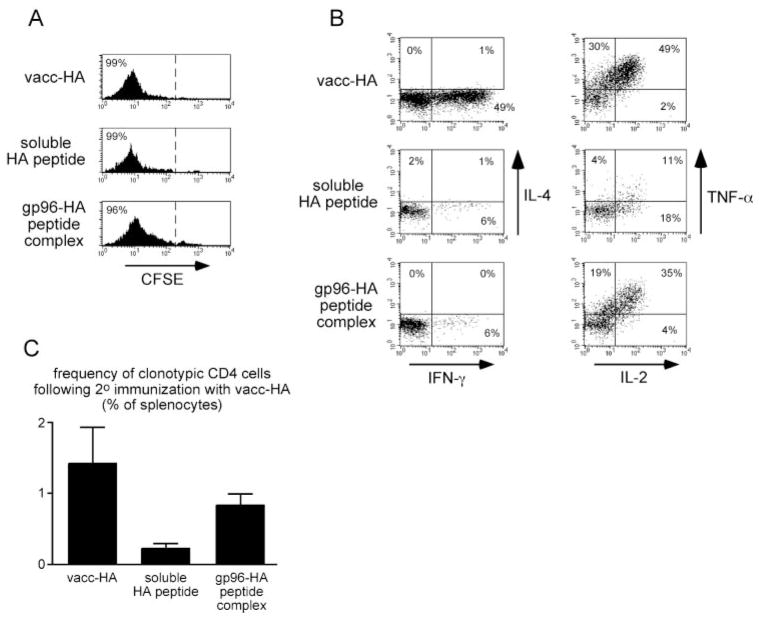
The ability of mature T lymphocytes to develop effector capacity after encounter with cognate Ag is generally dependent upon inflammatory signals associated with infection that induce dendritic cell activation/maturation. These inflammatory signals can derive directly from pathogens or can be expressed by host cells in response to infection. Heat shock proteins (HSPs) are a class of host-derived inflammatory mediators that perform the dual function of both chaperoning MHC class I-restricted epitopes into the cross-presentation pathway of DCs and inducing the activation/maturation of these DCs to allow priming of cognate CD8(+) T cell effector responses. Although the ability of HSPs to elicit effector CD8 cell responses has been well established, their potential to prime CD4 cell effector responses has been relatively unexplored. In the current study we compared the ability of the endoplasmic reticulum-resident HSP gp96 to prime CD4 vs CD8 cells using TCR transgenic adoptive transfer systems and soluble gp96-peptide complexes. As expected, gp96 facilitated the cross-presentation of a class I-restricted peptide and priming of effector function in cognate CD8 cells. Interestingly, gp96 also facilitated the in vivo presentation of a class II-restricted peptide; however, the resulting CD4 cell response did not involve the development of effector function. Taken together, these data suggest that gp96 is an inflammatory mediator that selectively primes CD8 cell effector function.
Figures
References
-
- Matzinger P. Tolerance, danger, and the extended family. Annu Rev Immunol. 1994;12:991. - PubMed
-
- Jenkins MK, Khoruts A, Ingulli E, Mueller DL, McSorley SJ, Reinhardt RL, Itano A, Pape KA. In vivo activation of antigen-specific CD4 T cells. Annu Rev Immunol. 2001;19:23. - PubMed
-
- Janeway CA, Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197. - PubMed
-
- Vella AT, McCormack JE, Linsley PS, Kappler JW, Marrack P. Lipopolysaccharide interferes with the induction of peripheral T cell death. Immunity. 1995;2:261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
