

doi: 10.1101/gad.1184704.
Epub 2004 May 6.
A combined computational-experimental approach predicts human microRNA targets
Affiliations
- PMID: 15131085
- PMCID: PMC415641
- DOI: 10.1101/gad.1184704
Item in Clipboard
A combined computational-experimental approach predicts human microRNA targets
Genes Dev.
.
Abstract
A new paradigm of gene expression regulation has emerged recently with the discovery of microRNAs (miRNAs). Most, if not all, miRNAs are thought to control gene expression, mostly by base pairing with miRNA-recognition elements (MREs) found in their messenger RNA (mRNA) targets. Although a large number of human miRNAs have been reported, many of their mRNA targets remain unknown. Here we used a combined bioinformatics and experimental approach to identify important rules governing miRNA-MRE recognition that allow prediction of human miRNA targets. We describe a computational program, "DIANA-microT", that identifies mRNA targets for animal miRNAs and predicts mRNA targets, bearing single MREs, for human and mouse miRNAs.
Figures
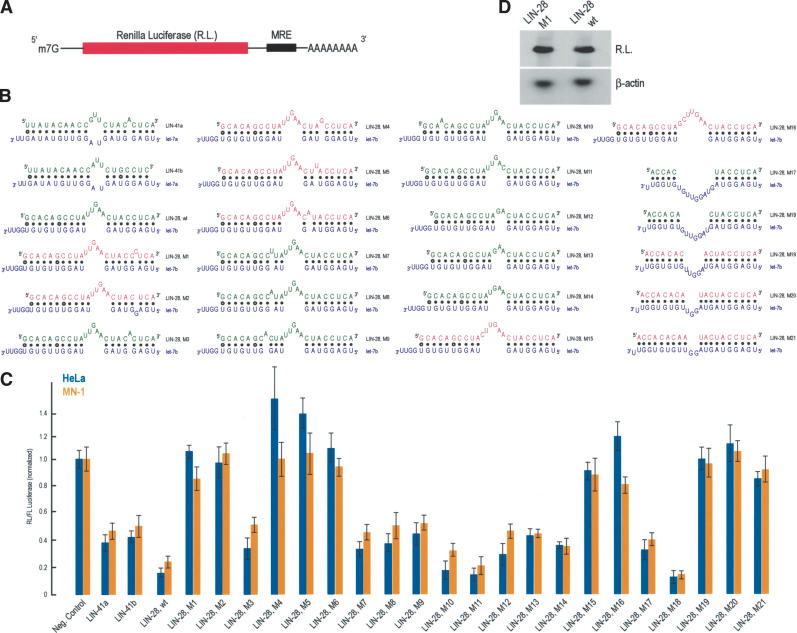
Experimental verification of a predicted miRNA recognition element (MRE) and deduction of the miRNA binding rules. (A) Schematic representation of the reporter construct. (red) Coding region. (B) Potential base pairing between predicted MREs derived from the indicated mRNAs and their cognate miRNAs (blue). (wt) Wild-type sequence. MRE sequences that repressed the expression of luciferase are shown in green; the sequences that that did not are shown in red. (C) HeLa human cells (blue bars) or MN-1 mouse cells (orange bars) were cotransfected with Renilla luciferase (RL) constructs bearing the indicated MREs in the 3′-UTR, along with firefly luciferase (FL). Results shown are average values (with standard deviations) of normalized RL/FL activities obtained from six separate experiments. (D) HeLa cells were transfected with indicated constructs, total RNA was isolated, and RL and β-actin (as a normalization control) mRNAs were visualized with Northern blots.
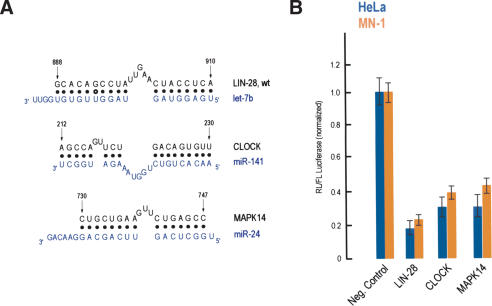
Predicted miRNA targets. (A) Potential base pairing between predicted MREs derived from the indicated mRNAs (black) and their cognate miRNAs (blue). Numbers refer to nucleotide positions after the stop codon, based on the human mRNAs. (B) HeLa (blue bars) or MN-1 (orange bars) cells were cotransfected with Renilla luciferase (RL) constructs bearing the indicated MREs in the 3′-UTR, along with firefly luciferase (FL). Results shown are average values (with standard deviations) of normalized RL/FL activities obtained from six separate experiments.

miRNA:MRE configurations that do not repress translation. (A) HeLa cells were cotransfected with Renilla luciferase (RL) constructs bearing the indicated MREs in the 3′-UTR, along with firefly luciferase (FL). (B) Results shown are average values (with standard deviations) of normalized RL/FL activities obtained from three separate experiments.
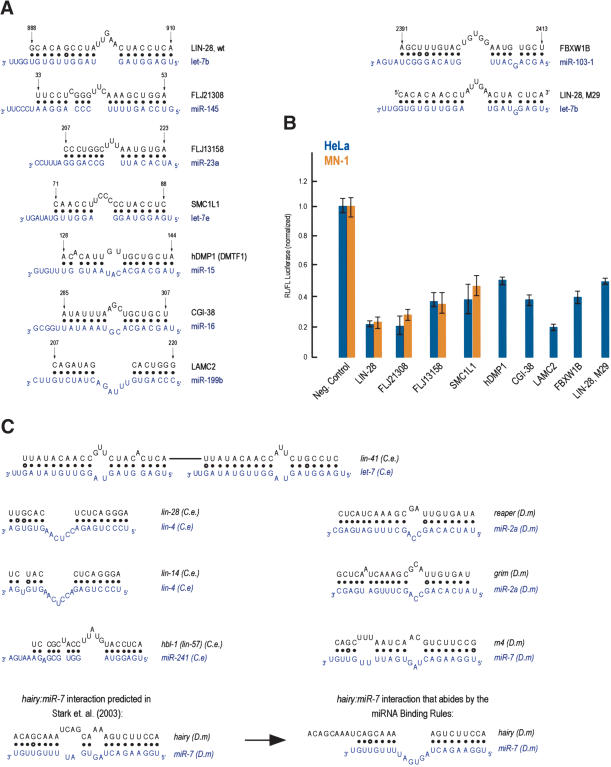
(A,B) Additional, predicted miRNA targets. Details as in the legend for Figure 3. The LIN-28, M29:let-7b was designed to mimic the binding characteristics of FBXWIB:miR-103 (which allows for a symmetrically placed single nucleotide bulge of the miRNA at the proximal region of the miRNA:MRE binding). (C) miRNA:MRE (target mRNA) interactions from C. elegans (C.e) or Drosophila melanogaster (D.m) targets that abide by the miRNA binding rules.
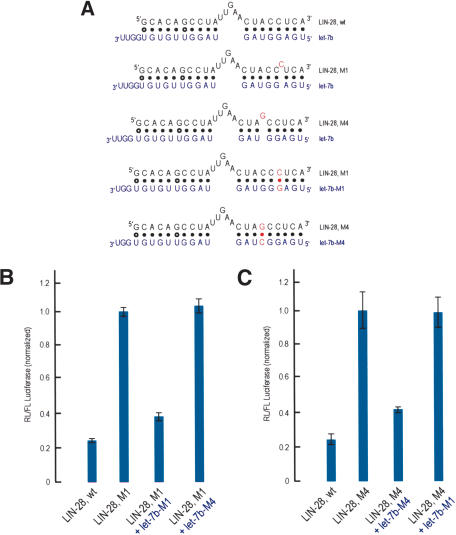
Restoration of miRNA-mediated translational repression of LIN-28 mutant MREs by synthetic let-7b siRNAs that carry compensatory mutations. (A) Potential base pairing between LIN-28 MREs (black) and endogenous let-7b or synthetic let-7b siRNAs (blue). Mutated nucleotides are shown in red. (B,C) HeLa cells were cotransfected with Renilla luciferase (RL) constructs bearing the indicated MREs in the 3′-UTR, along with firefly luciferase (FL) and with or without the indicated synthetic siRNAs (30 nM). Results shown are average values (with standard deviations) of normalized RL/FL activities obtained from three separate experiments.
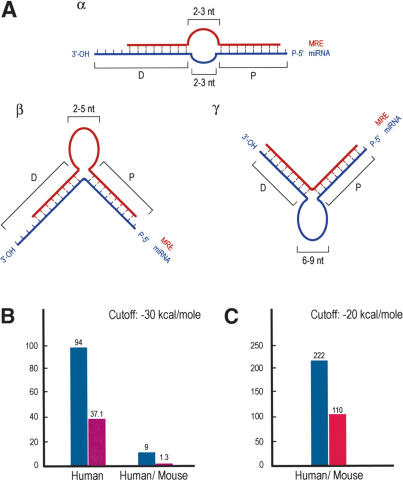
MicroRNA Binding Rules and Statistics. (A) Schematic representation of miRNA:MRE (target mRNA) bindings (miRNA binding rules). (Blue) miRNAs; (red) MRE; (P) proximal (relative to 5′-end of miRNA) region of miRNA:MRE binding; (D) distal region of binding. (Panel α) Loop, length (on each sequence) = 2–3 nt. (Panel β) Single MRE central bulge, length = 2–5 nt. (Panel γ) Single miRNA central bulge, length = 6–9 nt. Proximal binding characteristics are ≥7-nt base pairing between miRNA and MRE; the 5′-most nucleotide of the miRNA may or may not base pair with MRE; one symmetric single nucleotide bulge allowed (i.e., the single nucleotide bulge is surrounded by an equal number of base-paired nucleotides). Distal binding characteristics are ≥5-nt base pairing between miRNA and MRE; nucleotide bulges allowed. The last (toward the 3′ end) nucleotides of the miRNA may or may not base pair with the MRE. (B) Hits between 10 human miRNAs (blue bar) or shuffled RNAs (purple bar) and the 3′-UTR database of annotated human mRNAs or the conserved human/mouse 3′-UTR database (initial analysis). (C) Hits between 94 human miRNAs (blue bar) or shuffled controls (with the same compositional properties as the authentic miRNAs; red bar) and the conserved human/mouse 3′-UTR database extracted using EnsMart.
Similar articles
-
STarMir Tools for Prediction of microRNA Binding Sites.Methods Mol Biol. 2016;1490:73-82. doi: 10.1007/978-1-4939-6433-8_6. Methods Mol Biol. 2016. PMID: 27665594 Free PMC article.
-
Progress in miRNA target prediction and identification.Sci China C Life Sci. 2009 Dec;52(12):1123-30. doi: 10.1007/s11427-009-0159-4. Epub 2009 Dec 17. Sci China C Life Sci. 2009. PMID: 20016969 Review.
-
Analysis of microRNA-target interactions by a target structure based hybridization model.Pac Symp Biocomput. 2008:64-74. Pac Symp Biocomput. 2008. PMID: 18232104
-
Prediction and verification of microRNA targets by MovingTargets, a highly adaptable prediction method.BMC Genomics. 2005 Jun 8;6:88. doi: 10.1186/1471-2164-6-88. BMC Genomics. 2005. PMID: 15943864 Free PMC article.
-
Prediction of human microRNA targets.Methods Mol Biol. 2006;342:101-13. doi: 10.1385/1-59745-123-1:101. Methods Mol Biol. 2006. PMID: 16957370 Review.
Cited by
-
MicroRNA degeneracy and pluripotentiality within a Lavallière-tie architecture confers robustness to gene expression networks.Cell Mol Life Sci. 2016 Aug;73(15):2821-7. doi: 10.1007/s00018-016-2186-1. Epub 2016 Apr 1. Cell Mol Life Sci. 2016. PMID: 27038488 Free PMC article.
-
The Role of miR-212 and iNOS in Alcohol-Induced Intestinal Barrier Dysfunction and Steatohepatitis.Alcohol Clin Exp Res. 2015 Sep;39(9):1632-41. doi: 10.1111/acer.12813. Epub 2015 Jul 24. Alcohol Clin Exp Res. 2015. PMID: 26207424 Free PMC article.
-
MicroRNA: A Bridge from H. pylori Infection to Gastritis and Gastric Cancer Development.Front Genet. 2012 Dec 14;3:294. doi: 10.3389/fgene.2012.00294. eCollection 2012. Front Genet. 2012. PMID: 23248648 Free PMC article.
-
MicroRNA-93 may control vascular endothelial growth factor A in circulating peripheral blood mononuclear cells in acute Kawasaki disease.Pediatr Res. 2016 Sep;80(3):425-32. doi: 10.1038/pr.2016.93. Epub 2016 Apr 18. Pediatr Res. 2016. PMID: 27089500
-
Predicting kissing interactions in microRNA-target complex and assessment of microRNA activity.Nucleic Acids Res. 2012 May;40(10):4681-90. doi: 10.1093/nar/gks052. Epub 2012 Feb 3. Nucleic Acids Res. 2012. PMID: 22307238 Free PMC article.
References
-
- Abrahante J.E., Daul, A.L., Li, M., Volk, M.L., Tennessen, J.M., Miller, E.A., and Rougvie, A.E. 2003. The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by microRNAs. Dev. Cell 4: 625–637. - PubMed
-
- Ambros V., Lee, R.C., Lavanway, A., Williams, P.T., and Jewell, D. 2003b. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr. Biol. 13: 807–818. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases