

Nitrogen supply influences herbivore-induced direct and indirect defenses and transcriptional responses in Nicotiana attenuata
- PMID: 15133153
- PMCID: PMC429401
- DOI: 10.1104/pp.104.040360
Nitrogen supply influences herbivore-induced direct and indirect defenses and transcriptional responses in Nicotiana attenuata
Abstract
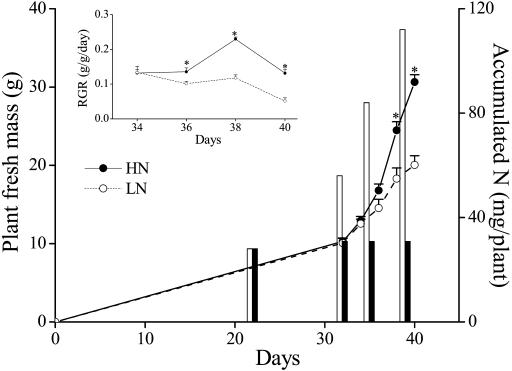
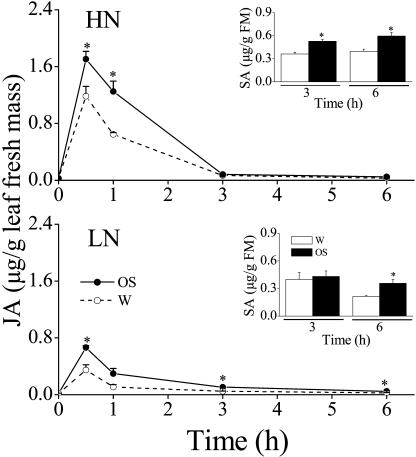
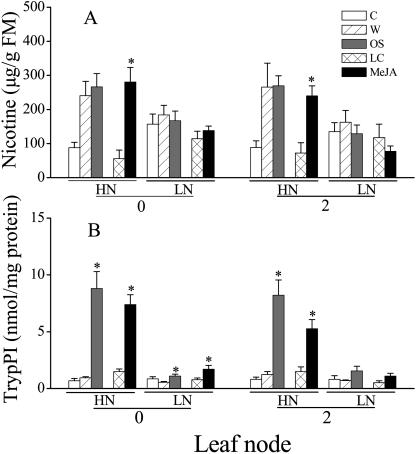
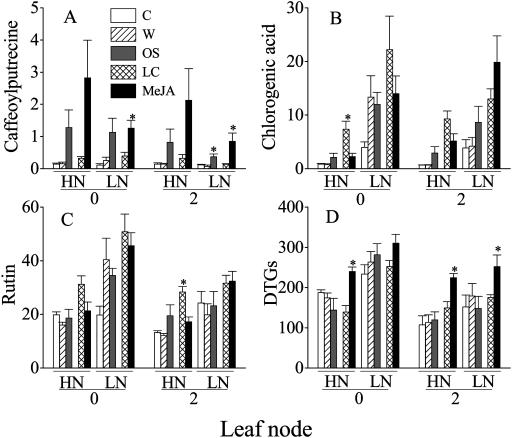
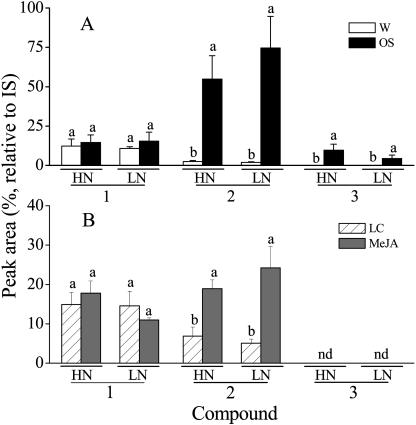
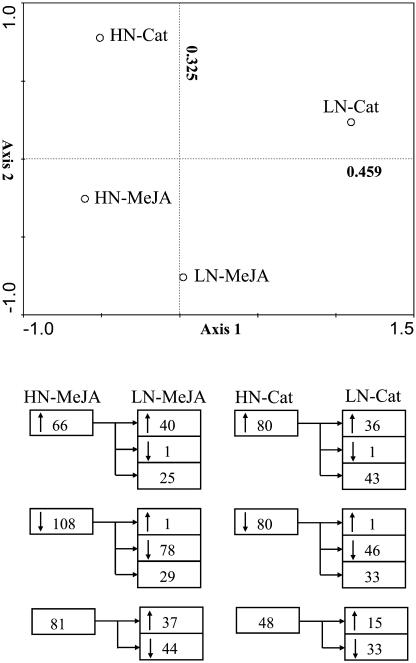
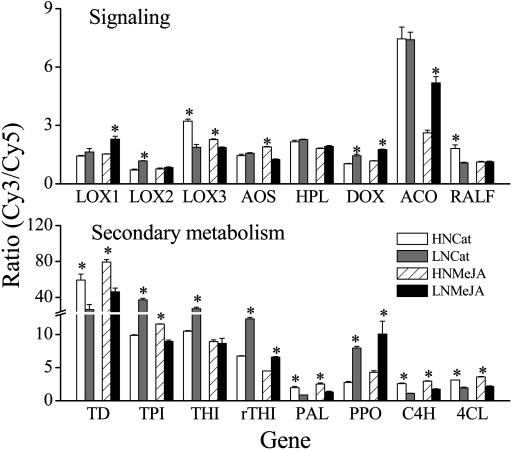
Although nitrogen (N) availability is known to alter constitutive resistance against herbivores, its influence on herbivore-induced responses, including signaling pathways, transcriptional signatures, and the subsequently elicited chemical defenses is poorly understood. We used the native tobacco, Nicotiana attenuata, which germinates in the postfire environment and copes with large changes in soil N during postfire succession, to compare a suite of Manduca sexta- and elicitor-induced responses in plants grown under high- and low-N (LN) supply rates. LN supply decreased relative growth rates and biomass by 35% at 40 d compared to high-N plants; furthermore, it also attenuated (by 39 and 60%) the elicitor-induced jasmonate and salicylate bursts, two N-intensive direct defenses (nicotine and trypsin proteinase inhibitors, albeit by different mechanisms), and carbon-containing nonvolatile defenses (rutin, chlorogenic acid, and diterpene glycosides), but did not affect the induced release of volatiles (cis-alpha-bergamotene and germacrene A), which function as indirect defenses. M. sexta and methyl jasmonate-induced transcriptional responses measured with a microarray enriched in herbivore-induced genes were also substantially reduced in plants grown under LN supply rates. In M. sexta-attacked LN plants, only 36 (45%) up-regulated and 46 (58%) down-regulated genes showed the same regulation as those in attacked high-N plants. However, transcriptional responses frequently directly countered the observed metabolic changes. Changes in a leaf's sensitivity to elicitation, an attacked leaf's waning ability to export oxylipin wound signals, and/or resource limitations in LN plants can account for the observed results, underscoring the conclusion that defense activation is a resource-intensive response.
Figures
References
-
- Baldwin IT (1999) Inducible nicotine production in native Nicotiana as an example of adaptive phenotypic plasticity. J Chem Ecol 25: 3–30
-
- Baldwin IT, Kessler A, Halitschke R (2002) Volatile signaling in plant-plant-herbivore interactions: What is real? Curr Opin Plant Biol 5: 351–354 - PubMed
-
- Baldwin IT, Staszak-Kozinski L, Davidson R (1994) Up in smoke: I. Smoke-derived germination cues for the postfire annual Nicotiana attenuata Torr. Ex Watson J Chem Ecol 20: 2345–2371 - PubMed
-
- Bryant JP, Chapin FS III, Klein DR (1983) Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40: 357–368
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
