

Neural induction in Xenopus: requirement for ectodermal and endomesodermal signals via Chordin, Noggin, beta-Catenin, and Cerberus
- PMID: 15138495
- PMCID: PMC406387
- DOI: 10.1371/journal.pbio.0020092
Neural induction in Xenopus: requirement for ectodermal and endomesodermal signals via Chordin, Noggin, beta-Catenin, and Cerberus
Abstract
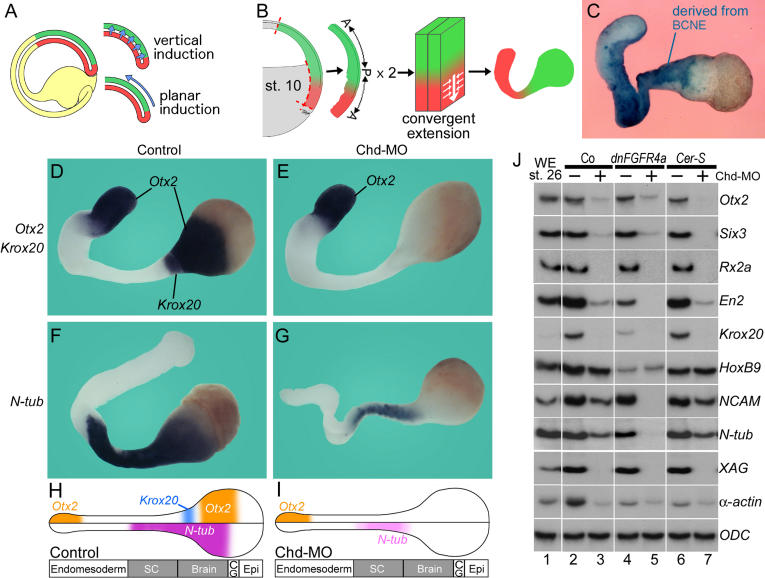
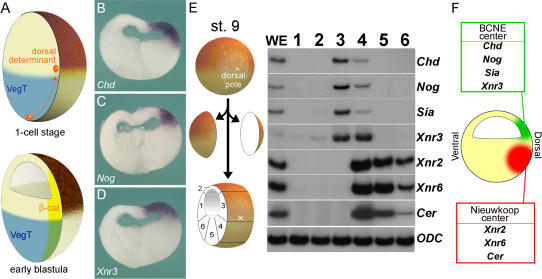
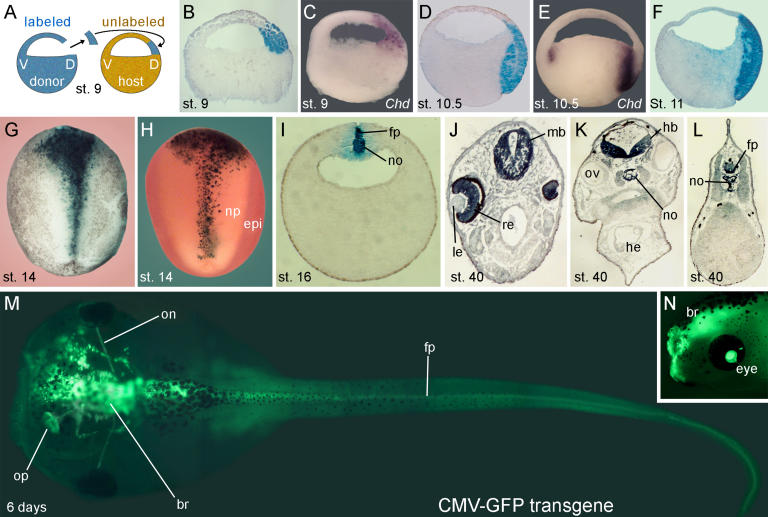
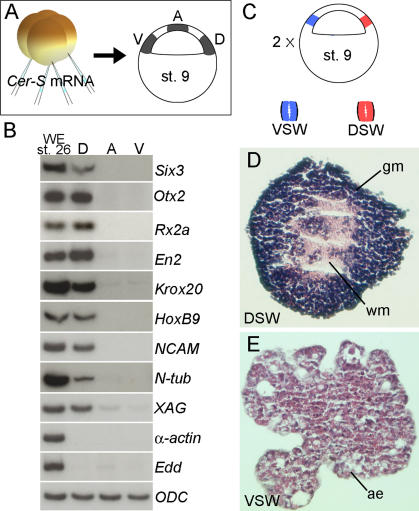
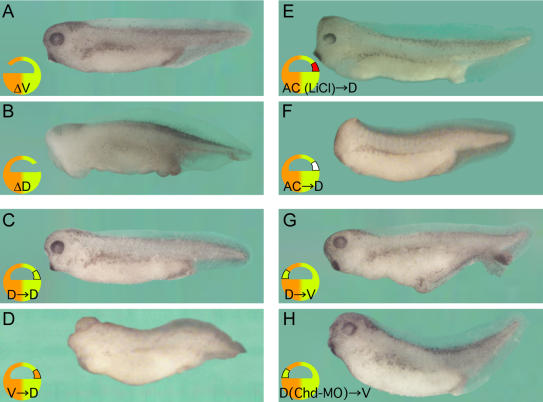
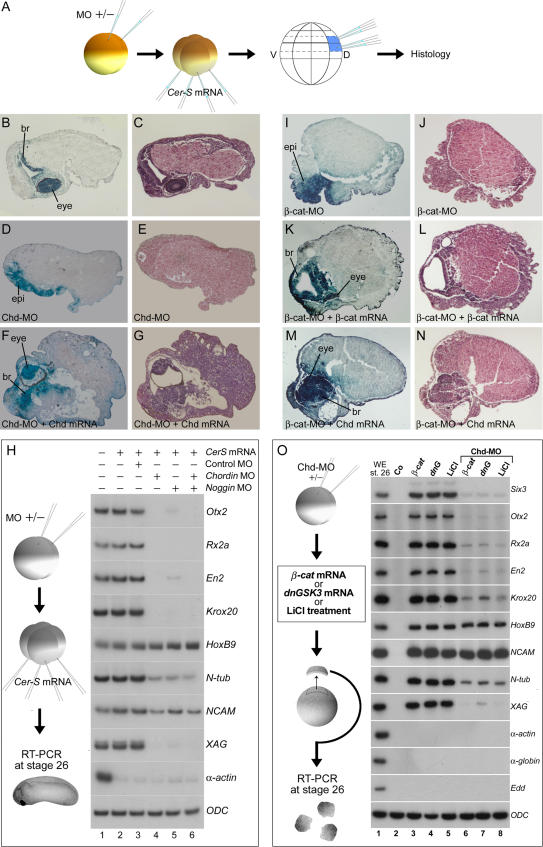
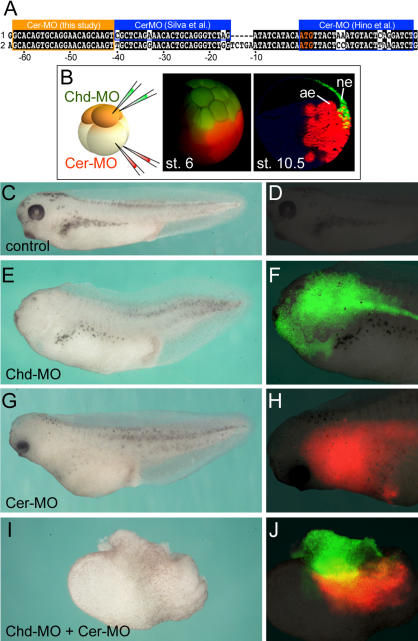
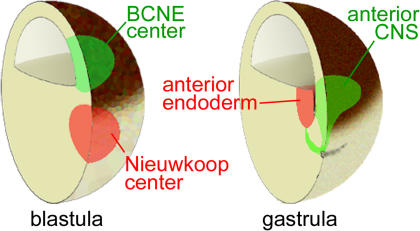
The origin of the signals that induce the differentiation of the central nervous system (CNS) is a long-standing question in vertebrate embryology. Here we show that Xenopus neural induction starts earlier than previously thought, at the blastula stage, and requires the combined activity of two distinct signaling centers. One is the well-known Nieuwkoop center, located in dorsal-vegetal cells, which expresses Nodal-related endomesodermal inducers. The other is a blastula Chordin- and Noggin-expressing (BCNE) center located in dorsal animal cells that contains both prospective neuroectoderm and Spemann organizer precursor cells. Both centers are downstream of the early beta-Catenin signal. Molecular analyses demonstrated that the BCNE center was distinct from the Nieuwkoop center, and that the Nieuwkoop center expressed the secreted protein Cerberus (Cer). We found that explanted blastula dorsal animal cap cells that have not yet contacted a mesodermal substratum can, when cultured in saline solution, express definitive neural markers and differentiate histologically into CNS tissue. Transplantation experiments showed that the BCNE region was required for brain formation, even though it lacked CNS-inducing activity when transplanted ventrally. Cell-lineage studies demonstrated that BCNE cells give rise to a large part of the brain and retina and, in more posterior regions of the embryo, to floor plate and notochord. Loss-of-function experiments with antisense morpholino oligos (MO) showed that the CNS that forms in mesoderm-less Xenopus embryos (generated by injection with Cerberus-Short [CerS] mRNA) required Chordin (Chd), Noggin (Nog), and their upstream regulator beta-Catenin. When mesoderm involution was prevented in dorsal marginal-zone explants, the anterior neural tissue formed in ectoderm was derived from BCNE cells and had a complete requirement for Chd. By injecting Chd morpholino oligos (Chd-MO) into prospective neuroectoderm and Cerberus morpholino oligos (Cer-MO) into prospective endomesoderm at the 8-cell stage, we showed that both layers cooperate in CNS formation. The results suggest a model for neural induction in Xenopus in which an early blastula beta-Catenin signal predisposes the prospective neuroectoderm to neural induction by endomesodermal signals emanating from Spemann's organizer.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
Similar articles
-
Neural induction in the absence of mesoderm: beta-catenin-dependent expression of secreted BMP antagonists at the blastula stage in Xenopus.Dev Biol. 2001 Jun 1;234(1):161-73. doi: 10.1006/dbio.2001.0258. Dev Biol. 2001. PMID: 11356027 Free PMC article.
-
Double assurance in the induction of axial development by egg dorsal determinants in Xenopus embryos.Proc Natl Acad Sci U S A. 2025 Feb 18;122(7):e2421772122. doi: 10.1073/pnas.2421772122. Epub 2025 Feb 10. Proc Natl Acad Sci U S A. 2025. PMID: 39928870 Free PMC article.
-
Dorsal-ventral patterning and neural induction in Xenopus embryos.Annu Rev Cell Dev Biol. 2004;20:285-308. doi: 10.1146/annurev.cellbio.20.011403.154124. Annu Rev Cell Dev Biol. 2004. PMID: 15473842 Free PMC article. Review.
-
Expression of Siamois and Twin in the blastula Chordin/Noggin signaling center is required for brain formation in Xenopus laevis embryos.Mech Dev. 2008 Jan-Feb;125(1-2):58-66. doi: 10.1016/j.mod.2007.10.005. Epub 2007 Oct 12. Mech Dev. 2008. PMID: 18036787 Free PMC article.
-
Pluripotent cells (stem cells) and their determination and differentiation in early vertebrate embryogenesis.Dev Growth Differ. 2001 Oct;43(5):469-502. doi: 10.1046/j.1440-169x.2001.00599.x. Dev Growth Differ. 2001. PMID: 11576166 Review.
Cited by
-
Nodal and <i>churchill1</i> position the expression of a notch ligand during <i>Xenopus</i> germ layer segregation.Life Sci Alliance. 2022 Sep 30;5(12):e202201693. doi: 10.26508/lsa.202201693. Life Sci Alliance. 2022. PMID: 36180230 Free PMC article.
-
Follistatin-like-1, a diffusible mesenchymal factor determines the fate of epithelium.Proc Natl Acad Sci U S A. 2010 Mar 9;107(10):4601-6. doi: 10.1073/pnas.0909501107. Epub 2010 Feb 22. Proc Natl Acad Sci U S A. 2010. PMID: 20176958 Free PMC article.
-
Role of BMP, FGF, calcium signaling, and Zic proteins in vertebrate neuroectodermal differentiation.Neurochem Res. 2011 Jul;36(7):1286-92. doi: 10.1007/s11064-011-0422-5. Epub 2011 Feb 19. Neurochem Res. 2011. PMID: 21336820 Free PMC article. Review.
-
Xenopus laevis as a Model Organism for the Study of Spinal Cord Formation, Development, Function and Regeneration.Front Neural Circuits. 2017 Nov 23;11:90. doi: 10.3389/fncir.2017.00090. eCollection 2017. Front Neural Circuits. 2017. PMID: 29218002 Free PMC article. Review.
-
ZC4H2 stabilizes Smads to enhance BMP signalling, which is involved in neural development in Xenopus.Open Biol. 2017 Aug;7(8):170122. doi: 10.1098/rsob.170122. Epub 2017 Aug 16. Open Biol. 2017. PMID: 28814648 Free PMC article.
References
-
- Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, et al. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature. 2000;403:658–661. - PubMed
-
- Bauer DV, Huang S, Moody SA. The cleavage stage origin of Spemann's organizer: Analysis of the movements of blastomere clones before and during gastrulation in Xenopus . Development. 1994;120:1179–1189. - PubMed
-
- Beddington RS, Robertson EJ. Axis development and early asymmetry in mammals. Cell. 2000;96:195–209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
