

Early myocardial function affects endocardial cushion development in zebrafish
- PMID: 15138499
- PMCID: PMC406391
- DOI: 10.1371/journal.pbio.0020129
Early myocardial function affects endocardial cushion development in zebrafish
Abstract
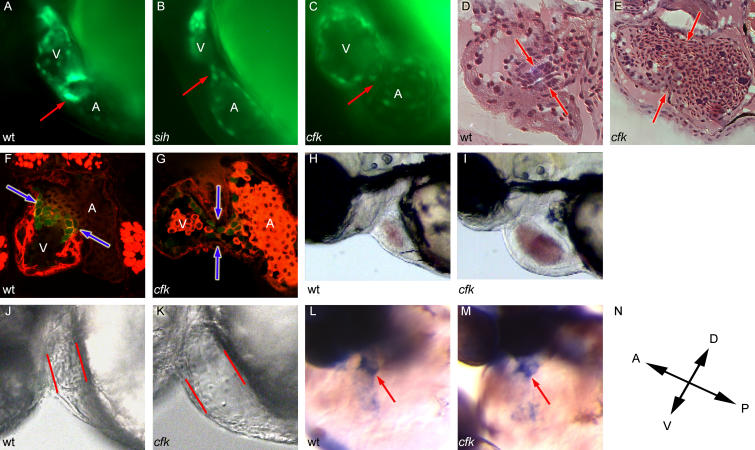
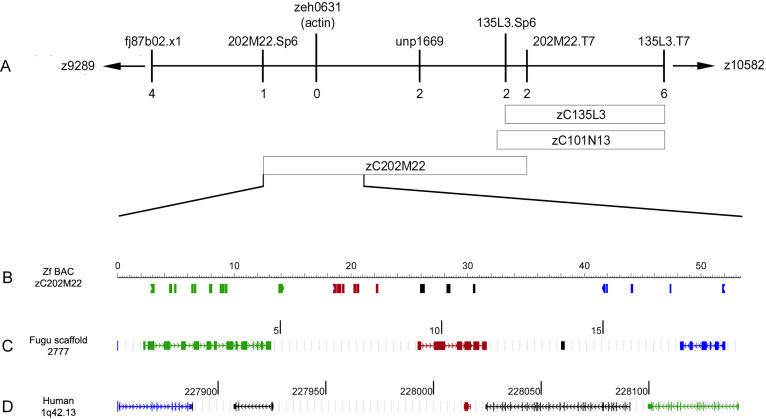
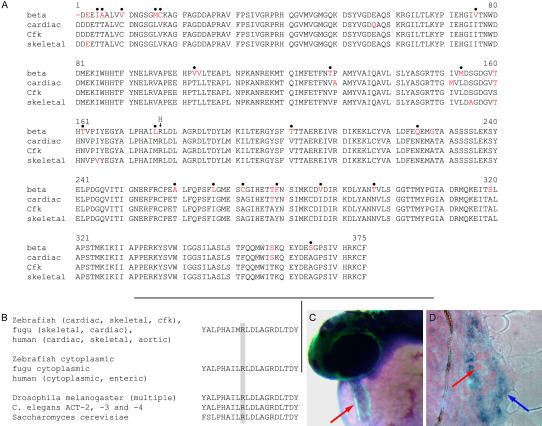
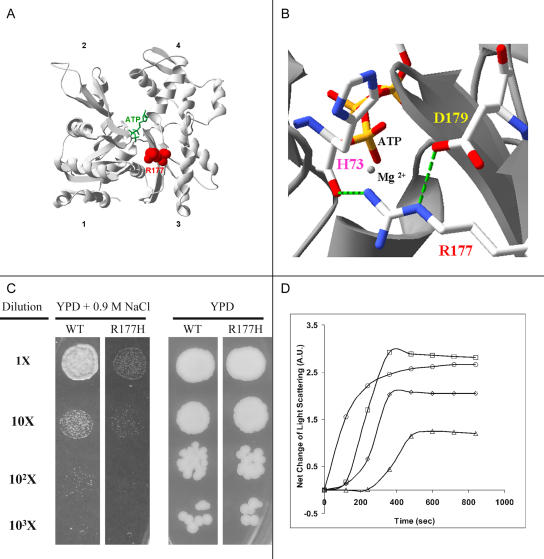
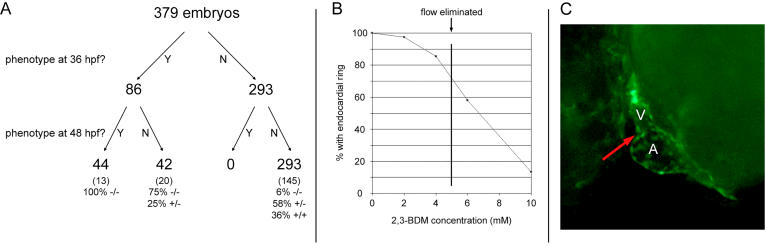
Function of the heart begins long before its formation is complete. Analyses in mouse and zebrafish have shown that myocardial function is not required for early steps of organogenesis, such as formation of the heart tube or chamber specification. However, whether myocardial function is required for later steps of cardiac development, such as endocardial cushion (EC) formation, has not been established. Recent technical advances and approaches have provided novel inroads toward the study of organogenesis, allowing us to examine the effects of both genetic and pharmacological perturbations of myocardial function on EC formation in zebrafish. To address whether myocardial function is required for EC formation, we examined silent heart (sih(-/-)) embryos, which lack a heartbeat due to mutation of cardiac troponin T (tnnt2), and observed that atrioventricular (AV) ECs do not form. Likewise, we determined that cushion formation is blocked in cardiofunk (cfk(-/-)) embryos, which exhibit cardiac dilation and no early blood flow. In order to further analyze the heart defects in cfk(-/-) embryos, we positionally cloned cfk and show that it encodes a novel sarcomeric actin expressed in the embryonic myocardium. The Cfk(s11) variant exhibits a change in a universally conserved residue (R177H). We show that in yeast this mutation negatively affects actin polymerization. Because the lack of cushion formation in sih- and cfk-mutant embryos could be due to reduced myocardial function and/or lack of blood flow, we approached this question pharmacologically and provide evidence that reduction in myocardial function is primarily responsible for the defect in cushion development. Our data demonstrate that early myocardial function is required for later steps of organogenesis and suggest that myocardial function, not endothelial shear stress, is the major epigenetic factor controlling late heart development. Based on these observations, we postulate that defects in cardiac morphogenesis may be secondary to mutations affecting early myocardial function, and that, in humans, mutations affecting embryonic myocardial function may be responsible for structural congenital heart disease.
Figures
References
-
- Alexander J, Stainier DY, Yelon D. Screening mosaic F1 females for mutations affecting zebrafish heart induction and patterning. Dev Genet. 1998;22:288–299. - PubMed
-
- Biben C, Weber R, Kesteven S, Stanley E, McDonald L, et al. Cardiac septal and valvular dysmorphogenesis in mice heterozygous for mutations in the homeobox gene Nkx2-5. Circ Res. 2000;87:888–895. - PubMed
-
- Bruneau BG, Nemer G, Schmitt JP, Charron F, Robitaille L, et al. A murine model of Holt–Oram syndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesis and disease. Cell. 2001;106:709–721. - PubMed
-
- Burge C, Karlin S. Prediction of complete gene structures in human genomic DNA. J Mol Biol. 1997;268:78–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
