

Multiple reference frames for motion in the primate cerebellum
- PMID: 15140919
- PMCID: PMC6729386
- DOI: 10.1523/JNEUROSCI.0109-04.2004
Multiple reference frames for motion in the primate cerebellum
Abstract
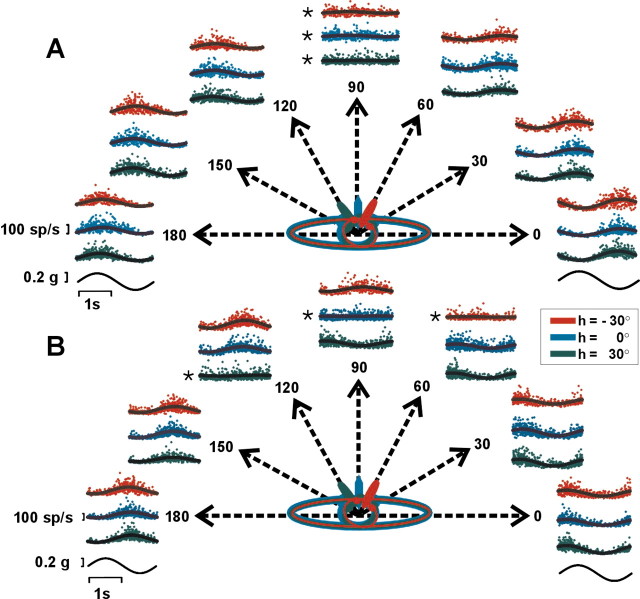
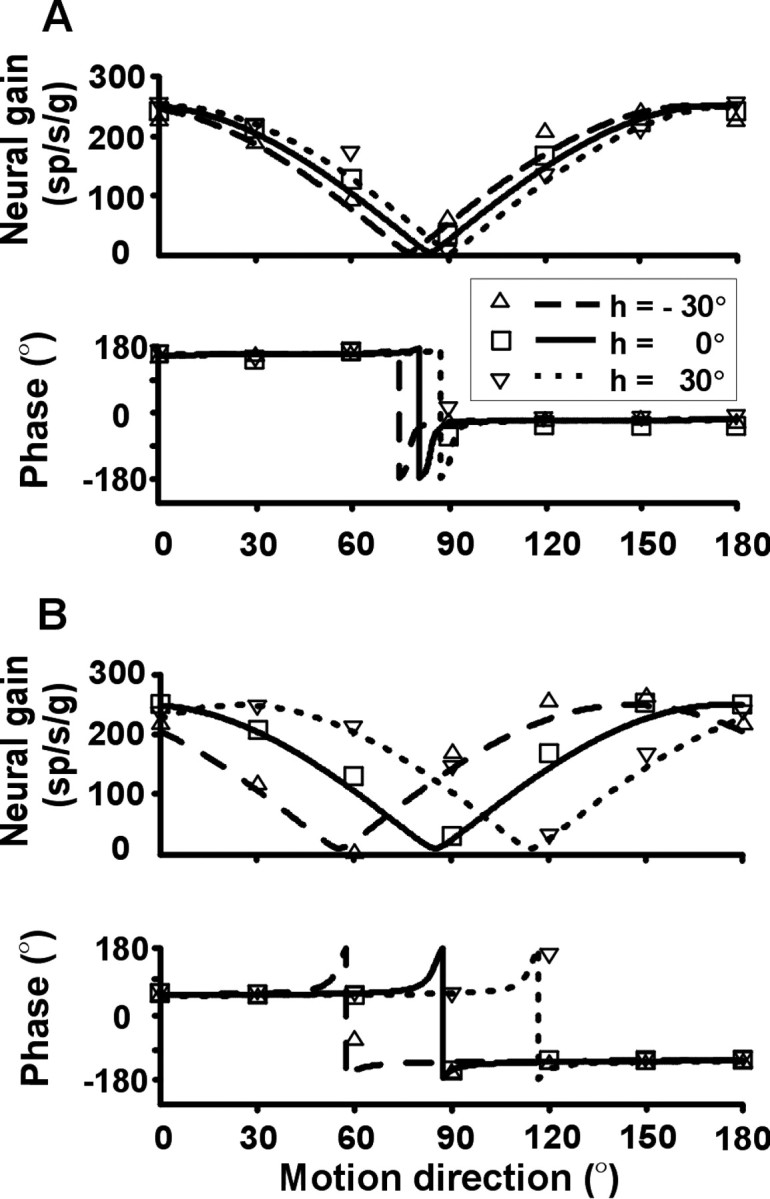
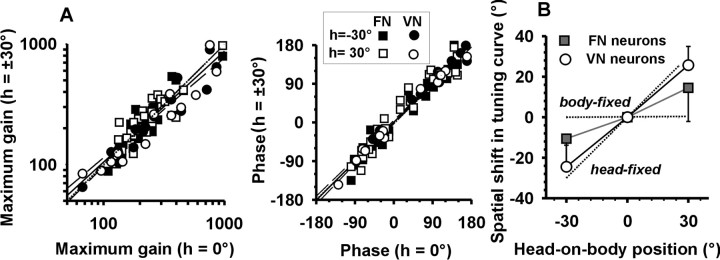
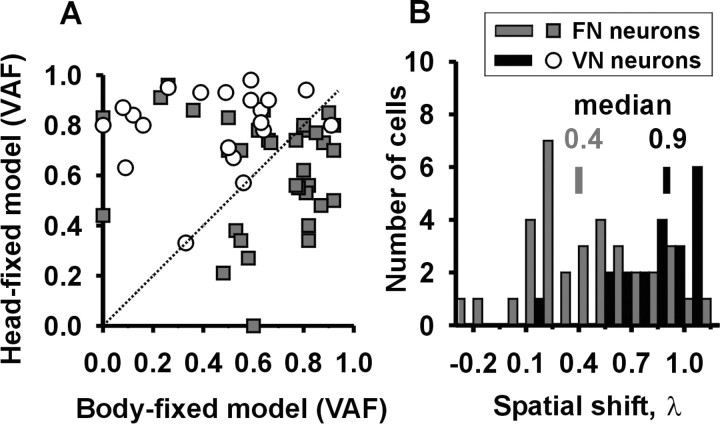
Knowledge of body motion through space is necessary for spatial orientation, self-motion perception, and postural control. Yet, sensory afferent signals may not directly provide such information to the brain. Because motion detected by the vestibular end organs is encoded in a head-fixed frame of reference, a coordinate transformation is thus required to encode body motion. In this study, we investigated whether cerebellar motion-sensitive neurons encode the translation of the body through space. We systematically changed both the direction of motion relative to the body and the static orientation of the head relative to the trunk. The activities of motion-sensitive neurons in the most medial of the deep cerebellar nuclei, the rostral fastigial nucleus, were compared with those in the brainstem vestibular nuclei. We found a distributed representation of reference frames for motion in the rostral fastigial nucleus, in contrast to cells in the vestibular nuclei, which primarily encoded motion in a head-fixed reference frame. This differential representation of motion-related information implies potential differences in the functional roles of these areas.
Figures
Similar articles
-
Properties of cerebellar fastigial neurons during translation, rotation, and eye movements.J Neurophysiol. 2005 Feb;93(2):853-63. doi: 10.1152/jn.00879.2004. Epub 2004 Sep 15. J Neurophysiol. 2005. PMID: 15371498
-
Role of Rostral Fastigial Neurons in Encoding a Body-Centered Representation of Translation in Three Dimensions.J Neurosci. 2018 Apr 4;38(14):3584-3602. doi: 10.1523/JNEUROSCI.2116-17.2018. Epub 2018 Feb 27. J Neurosci. 2018. PMID: 29487123 Free PMC article.
-
Multimodal integration in rostral fastigial nucleus provides an estimate of body movement.J Neurosci. 2009 Aug 26;29(34):10499-511. doi: 10.1523/JNEUROSCI.1937-09.2009. J Neurosci. 2009. PMID: 19710303 Free PMC article.
-
Gravity or translation: central processing of vestibular signals to detect motion or tilt.J Vestib Res. 2003;13(4-6):245-53. J Vestib Res. 2003. PMID: 15096668 Review.
-
Central vestibular system: vestibular nuclei and posterior cerebellum.Brain Res Bull. 2003 Jun 15;60(5-6):511-41. doi: 10.1016/s0361-9230(03)00055-8. Brain Res Bull. 2003. PMID: 12787870 Review.
Cited by
-
Proprioception and the predictive sensing of active self-motion.Curr Opin Physiol. 2021 Apr;20:29-38. doi: 10.1016/j.cophys.2020.12.001. Epub 2021 Jan 22. Curr Opin Physiol. 2021. PMID: 33954270 Free PMC article.
-
Optokinetic and vestibular responsiveness in the macaque rostral vestibular and fastigial nuclei.J Neurophysiol. 2009 Feb;101(2):714-20. doi: 10.1152/jn.90612.2008. Epub 2008 Dec 10. J Neurophysiol. 2009. PMID: 19073813 Free PMC article.
-
Multisensory integration in early vestibular processing in mice: the encoding of passive vs. active motion.J Neurophysiol. 2013 Dec;110(12):2704-17. doi: 10.1152/jn.01037.2012. Epub 2013 Oct 2. J Neurophysiol. 2013. PMID: 24089394 Free PMC article.
-
Cerebellum Lecture: the Cerebellar Nuclei-Core of the Cerebellum.Cerebellum. 2024 Apr;23(2):620-677. doi: 10.1007/s12311-022-01506-0. Epub 2023 Feb 13. Cerebellum. 2024. PMID: 36781689 Free PMC article. Review.
-
Does Inferior-Olive Hypersynchrony Affect Vestibular Heading Perception?Cerebellum. 2021 Oct;20(5):744-750. doi: 10.1007/s12311-020-01103-z. Cerebellum. 2021. PMID: 31939030
References
-
- Amino Y, Kyuhou S, Matsuzaki R, Gemba H (2001) Cerebello-thalamocortical projections to the posterior parietal cortex in the macaque monkey. Neurosci Lett 309: 29-32. - PubMed
-
- Anastasopoulos D, Mergner T (1982) Canal-neck interaction in vestibular nuclear neurons of the cat. Exp Brain Res 46: 269-280. - PubMed
-
- Angelaki DE (1991) Dynamic polarization vector of spatially tuned neurons. IEEE Trans Biomed Eng 38: 1053-1060. - PubMed
-
- Angelaki DE, Dickman JD (2000) Spatiotemporal processing of linear acceleration: primary afferent and central vestibular neuron responses. J Neurophysiol 84: 2113-2132. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources