

A Drosophila temperature-sensitive seizure mutant in phosphoglycerate kinase disrupts ATP generation and alters synaptic function
- PMID: 15140922
- PMCID: PMC6729406
- DOI: 10.1523/JNEUROSCI.0542-04.2004
A Drosophila temperature-sensitive seizure mutant in phosphoglycerate kinase disrupts ATP generation and alters synaptic function
Abstract
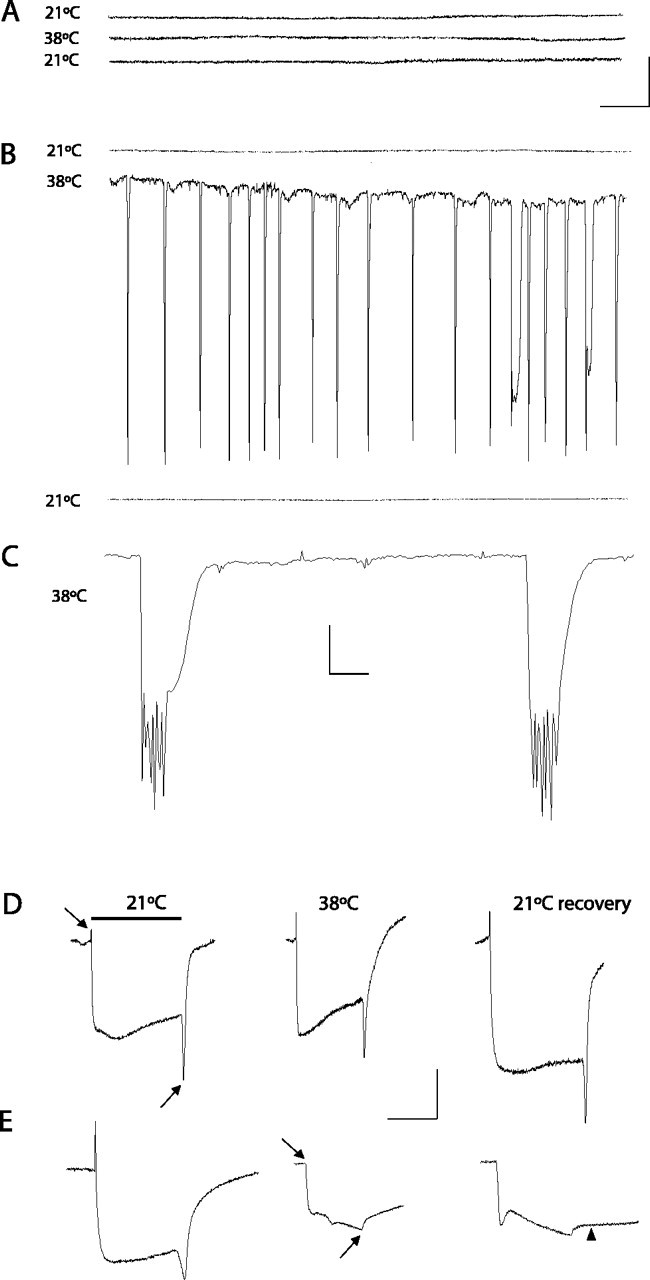
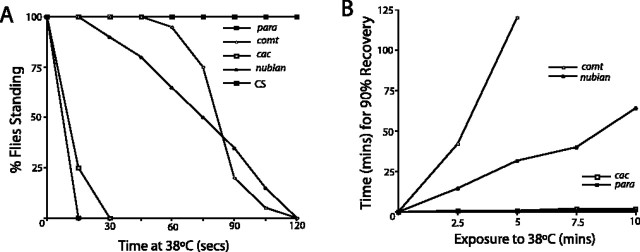
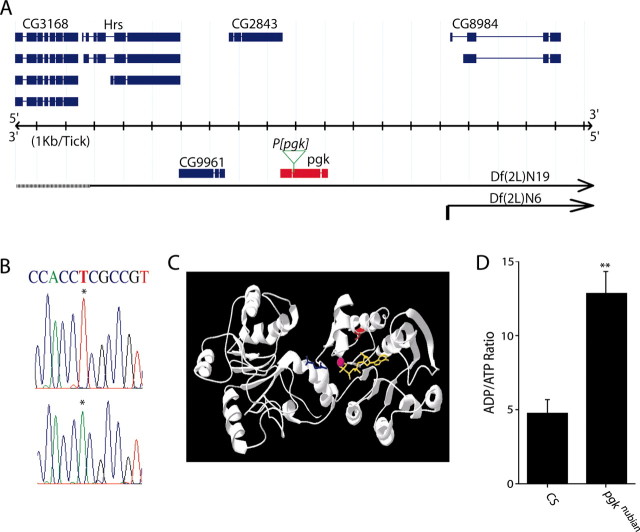
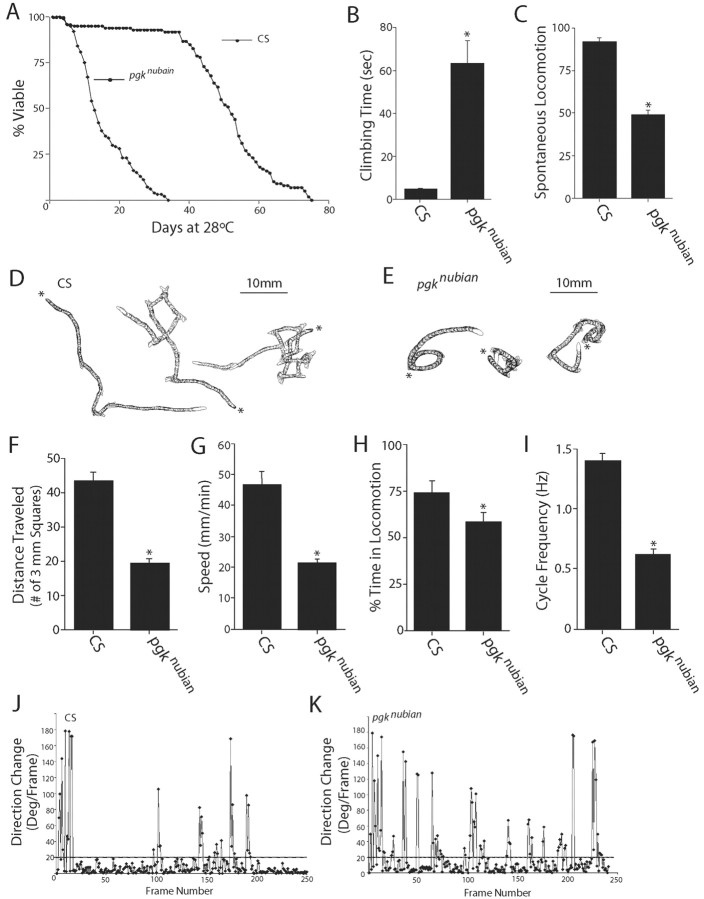
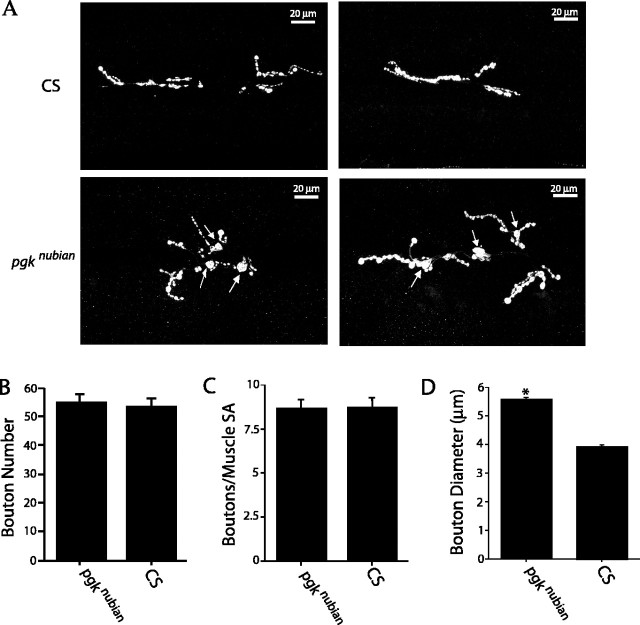
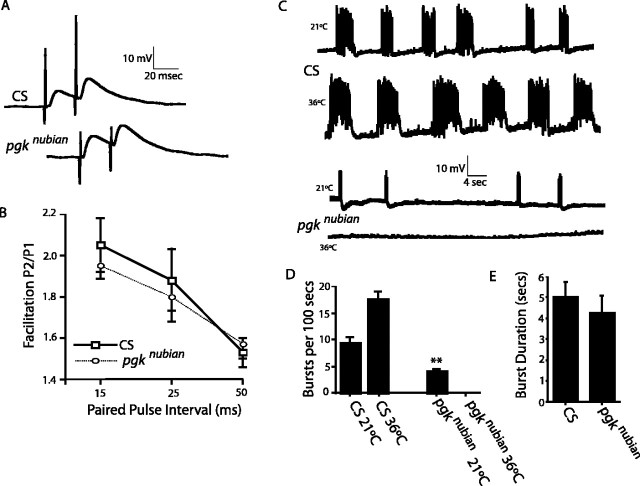
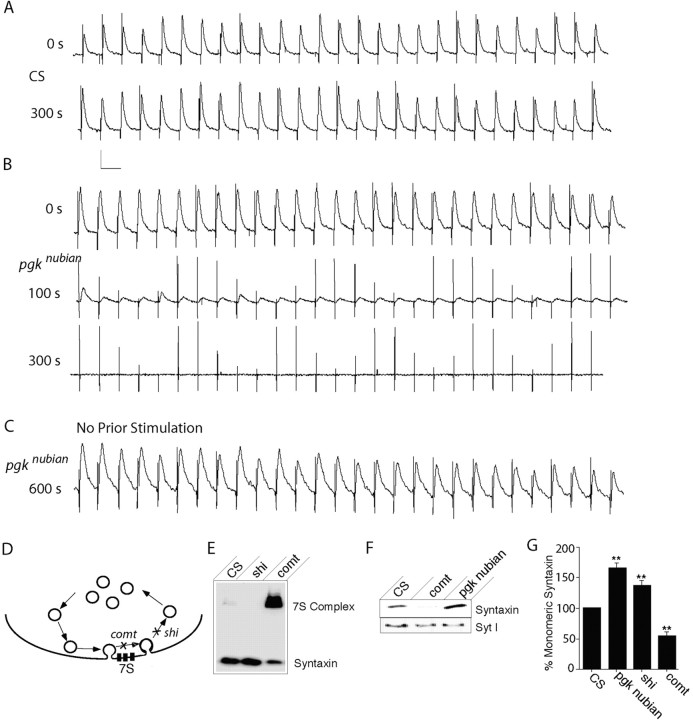
A novel paralytic mutant, nubian, was identified in a behavioral screen for conditional temperature-sensitive seizure mutants in Drosophila melanogaster. nubian mutants display reduced lifespan, abnormal motor behavior, altered synaptic structure, and defective neurotransmitter release. The nubian mutant disrupts phosphoglycerate kinase (PGK), an enzyme required for ATP generation in the terminal stage of the glycolytic pathway. Consistent with altered ATP generation in nubian animals, brain extracts show a threefold reduction in resting ATP levels compared with controls. Microarray analysis of nubian mutants reveals altered transcription of genes implicated in glucose and lipid metabolism. Disruption of ATP generation in nubian animals is accompanied by temperature-dependent defects in neuronal activity, with initial seizure activity, followed by an activity-dependent loss of synaptic transmission. nubian mutants also display structural defects at the synapse, with larger varicosity size but normal varicosity number, indicating that these synaptic parameters are regulated independently. Both exocytotic (NSF) and endocytotic (dynamin) ATPase/GTPase activity are required for normal synaptic transmission. Biochemical and physiological analyses indicate that synaptic defects in nubian animals are secondary to defective endocytosis, suggesting that endocytotic pathways may be generally more sensitive to altered ATP levels than those used for exocytosis. Alterations in ATP metabolism likely disrupt similar pathways in humans, because PGK deficiency is associated with mental retardation, seizures, and exercise intolerance. Given the behavioral similarities between disruptions of PGK function in Drosophila and humans, the analysis of nubian animals may reveal conserved neuronal responses associated with altered ATP generation within the brain.
Figures
Similar articles
-
Genetic interaction between shibire and comatose mutations in Drosophila suggest a role for snap-receptor complex assembly and disassembly for maintenance of synaptic vesicle cycling.Neurosci Lett. 2001 Sep 21;311(1):21-4. doi: 10.1016/s0304-3940(01)02125-5. Neurosci Lett. 2001. PMID: 11585558
-
Dominant-negative NSF2 disrupts the structure and function of Drosophila neuromuscular synapses.J Neurobiol. 2002 Jun 15;51(4):261-71. doi: 10.1002/neu.10059. J Neurobiol. 2002. PMID: 12150502
-
NSF function in neurotransmitter release involves rearrangement of the SNARE complex downstream of synaptic vesicle docking.J Neurosci. 1998 Dec 15;18(24):10250-6. doi: 10.1523/JNEUROSCI.18-24-10250.1998. J Neurosci. 1998. PMID: 9852562 Free PMC article.
-
Genetic modifiers of comatose mutations in Drosophila: insights into neuronal NSF (N-ethylmaleimide-sensitive fusion factor) functions.J Neurogenet. 2012 Sep;26(3-4):348-59. doi: 10.3109/01677063.2012.697500. Epub 2012 Jul 20. J Neurogenet. 2012. PMID: 22817636 Review.
-
The synaptic vesicle cycle: exocytosis and endocytosis in Drosophila and C. elegans.Curr Opin Neurobiol. 2002 Oct;12(5):499-507. doi: 10.1016/s0959-4388(02)00360-4. Curr Opin Neurobiol. 2002. PMID: 12367628 Review.
Cited by
-
Structural and Genetic Studies Demonstrate Neurologic Dysfunction in Triosephosphate Isomerase Deficiency Is Associated with Impaired Synaptic Vesicle Dynamics.PLoS Genet. 2016 Mar 31;12(3):e1005941. doi: 10.1371/journal.pgen.1005941. eCollection 2016 Mar. PLoS Genet. 2016. PMID: 27031109 Free PMC article.
-
Characterization of the role of the Synaptotagmin family as calcium sensors in facilitation and asynchronous neurotransmitter release.Proc Natl Acad Sci U S A. 2007 Aug 28;104(35):14122-7. doi: 10.1073/pnas.0706711104. Epub 2007 Aug 20. Proc Natl Acad Sci U S A. 2007. PMID: 17709738 Free PMC article.
-
E2F regulation of the Phosphoglycerate kinase gene is functionally important in Drosophila development.Proc Natl Acad Sci U S A. 2023 Apr 11;120(15):e2220770120. doi: 10.1073/pnas.2220770120. Epub 2023 Apr 3. Proc Natl Acad Sci U S A. 2023. PMID: 37011211 Free PMC article.
-
A genetic locus for sensory epilepsy precipitated by contact with hot water maps to chromosome 9p24.3-p23.J Genet. 2018 Jun;97(2):391-398. J Genet. 2018. PMID: 29932058
-
Secretory Carrier Membrane Protein (SCAMP) deficiency influences behavior of adult flies.Front Cell Dev Biol. 2014 Nov 18;2:64. doi: 10.3389/fcell.2014.00064. eCollection 2014. Front Cell Dev Biol. 2014. PMID: 25478561 Free PMC article.
References
-
- Akasu T, Tsurusaki M, Shoji S (1996) Depletion of glucose causes presynaptic inhibition of neuronal transmission in the rat dorsolateral septal nucleus. Synapse 24: 125-134. - PubMed
-
- Atkinson NS, Robertson GA, Ganetzky B (1991) A component of calcium-activated potassium channels encoded by the Drosophila slo locus. Science 253: 551-555. - PubMed
-
- Bernstein BE, Michels PA, Hol WG (1997) Synergistic effects of substrate-induced conformational changes in phosphoglycerate kinase activation. Nature 385: 275-278. - PubMed
-
- Davis GW, Bezprozvanny I (2001) Maintaining the stability of neural function: a homeostatic hypothesis. Annu Rev Physiol 63: 847-869. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases