

Suppression of proinflammatory cytokine expression by herpes simplex virus type 1
- PMID: 15140986
- PMCID: PMC415838
- DOI: 10.1128/JVI.78.11.5883-5890.2004
Suppression of proinflammatory cytokine expression by herpes simplex virus type 1
Abstract
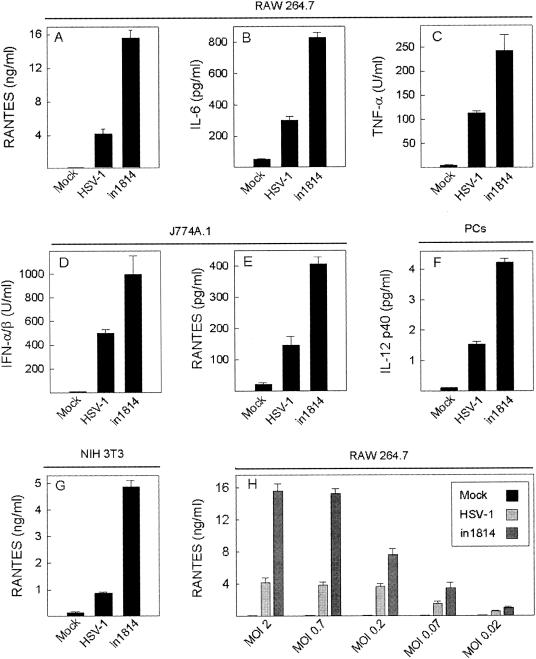
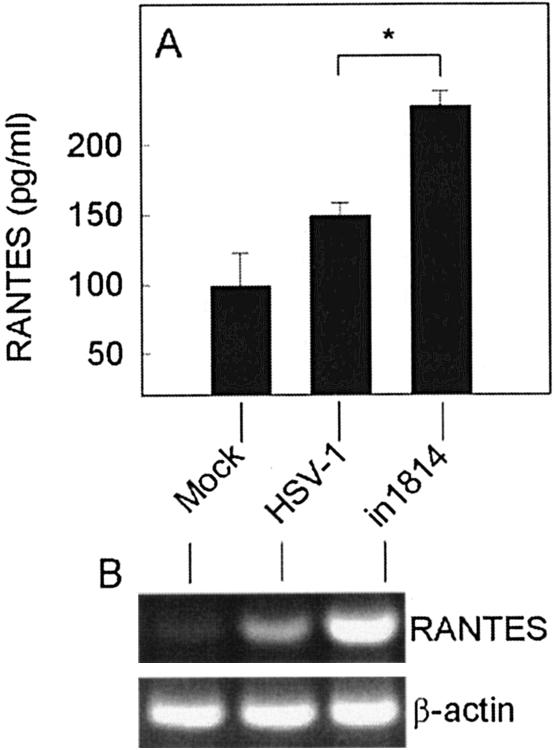
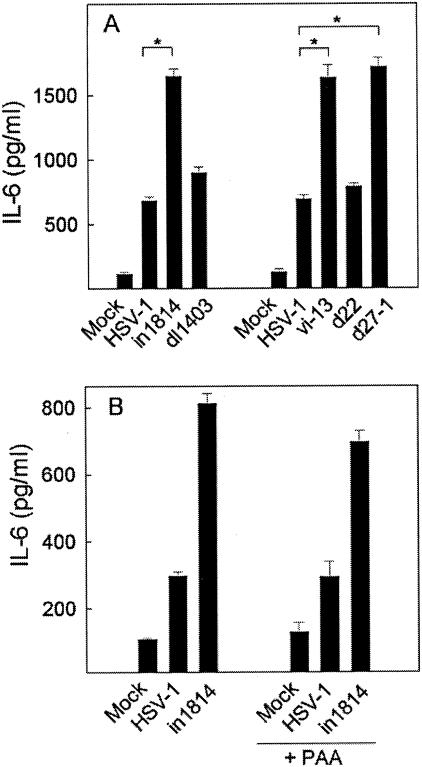
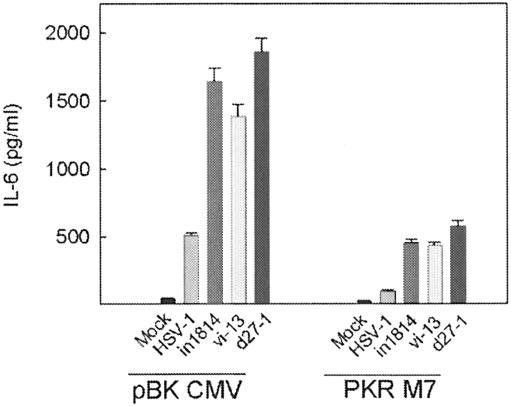
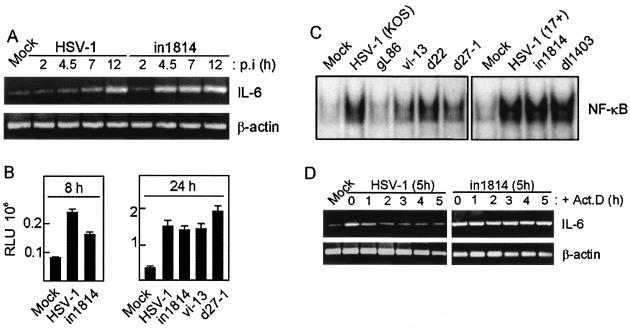
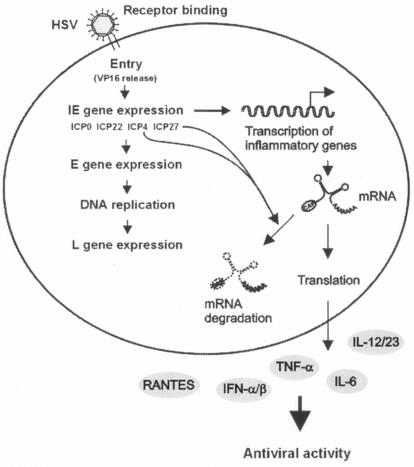
Viral immune evasion strategies are important for establishment and maintenance of infections. Many viruses are in possession of mechanisms to counteract the antiviral response raised by the infected host. Here we show that a herpes simplex virus type 1 (HSV-1) mutant lacking functional viral protein 16 (VP16)-a tegument protein promoting viral gene expression-induced significantly higher levels of proinflammatory cytokines than wild-type HSV-1. This was observed in several cell lines and primary murine macrophages, as well as in peritoneal cells harvested from mice infected in vivo. The enhanced ability to stimulate cytokine expression in the absence of VP16 was not mediated directly by VP16 but was dependent on the viral immediate-early genes for infected cell protein 4 (ICP4) and ICP27, which are expressed in a VP16-dependent manner during primary HSV infection. The virus appeared to target cellular factors other than interferon-induced double-stranded RNA-activated protein kinase R (PKR), since the virus mutants remained stronger inducers of cytokines in cells stably expressing a dominant-negative mutant form of PKR. Finally, mRNA stability assay revealed a significantly longer half-life for interleukin-6 mRNA after infection with the VP16 mutant than after infection with the wild-type virus. Thus, HSV is able to suppress expression of proinflammatory cytokines by decreasing the stability of mRNAs, thereby potentially impeding the antiviral host response to infection.
Figures
Similar articles
-
Virus-cell interactions regulating induction of tumor necrosis factor alpha production in macrophages infected with herpes simplex virus.J Virol. 2001 Nov;75(21):10170-8. doi: 10.1128/JVI.75.21.10170-10178.2001. J Virol. 2001. PMID: 11581385 Free PMC article.
-
Control of VP16 translation by the herpes simplex virus type 1 immediate-early protein ICP27.J Virol. 2005 Apr;79(7):4120-31. doi: 10.1128/JVI.79.7.4120-4131.2005. J Virol. 2005. PMID: 15767413 Free PMC article.
-
Spread and replication of and immune response to gamma134.5-negative herpes simplex virus type 1 vectors in BALB/c mice.J Virol. 2004 Dec;78(23):13139-52. doi: 10.1128/JVI.78.23.13139-13152.2004. J Virol. 2004. PMID: 15542666 Free PMC article.
-
[Research Advances in VP16 of the Herpes Virus].Bing Du Xue Bao. 2016 Nov;32(6):817-24. Bing Du Xue Bao. 2016. PMID: 30004657 Review. Chinese.
-
[Molecular mechanisms for suppression of interferon system by HSV-1 infection].Nihon Rinsho. 2006 Mar;64 Suppl 3:171-7. Nihon Rinsho. 2006. PMID: 16615462 Review. Japanese. No abstract available.
Cited by
-
Genomic analysis of host - Peste des petits ruminants vaccine viral transcriptome uncovers transcription factors modulating immune regulatory pathways.Vet Res. 2015 Feb 24;46:15. doi: 10.1186/s13567-015-0153-8. Vet Res. 2015. PMID: 25827022 Free PMC article.
-
Role of Bacterial Exopolysaccharides as Agents in Counteracting Immune Disorders Induced by Herpes Virus.Microorganisms. 2015 Aug 21;3(3):464-83. doi: 10.3390/microorganisms3030464. Microorganisms. 2015. PMID: 27682100 Free PMC article. Review.
-
Human cytomegalovirus immediate-early 2 protein IE86 blocks virus-induced chemokine expression.J Virol. 2006 Jan;80(2):920-8. doi: 10.1128/JVI.80.2.920-928.2006. J Virol. 2006. PMID: 16378994 Free PMC article.
-
Inhibition of herpes simplex-1 virus replication by 25-hydroxycholesterol and 27-hydroxycholesterol.Redox Biol. 2017 Aug;12:522-527. doi: 10.1016/j.redox.2017.03.016. Epub 2017 Mar 23. Redox Biol. 2017. PMID: 28359048 Free PMC article.
-
Vaccinium bracteatum Thunb Extract Inhibits HSV-1 Infection by Regulating ER Stress and Apoptosis.Antioxidants (Basel). 2022 Sep 8;11(9):1773. doi: 10.3390/antiox11091773. Antioxidants (Basel). 2022. PMID: 36139847 Free PMC article.
References
-
- Biron, C. A. 1999. Initial and innate responses to viral infections. Curr. Opin. Microbiol. 2:374-381. - PubMed
-
- Biron, C. A., K. S. Byron, and J. L. Sullivan. 1989. Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med. 320:1731-1735. - PubMed
-
- Campbell, M. E., J. W. Palfreyman, and C. M. Preston. 1984. Identification of herpes simplex virus DNA sequences which encode a trans-acting polypeptide responsible for stimulation of immediate early transcription. J. Mol. Biol. 25:1-19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
