

doi: 10.1128/JVI.78.11.6061-6066.2004.
Reverse genetic analysis of the transcription regulatory sequence of the coronavirus transmissible gastroenteritis virus
Affiliations
- PMID: 15141005
- PMCID: PMC415797
- DOI: 10.1128/JVI.78.11.6061-6066.2004
Item in Clipboard
Reverse genetic analysis of the transcription regulatory sequence of the coronavirus transmissible gastroenteritis virus
J Virol.
2004 Jun.
Abstract
Coronavirus discontinuous transcription uses a highly conserved sequence (CS) in the joining of leader and body RNAs. Using a full-length infectious construct of transmissable gastroenteritis virus, the present study demonstrates that subgenomic transcription is heavily influenced by upstream flanking sequences and supports a mechanism of transcription attenuation that is regulated in part by a larger domain composed of primarily upstream flanking sequences which select appropriately positioned CS elements for synthesis of subgenomic RNAs.
Figures
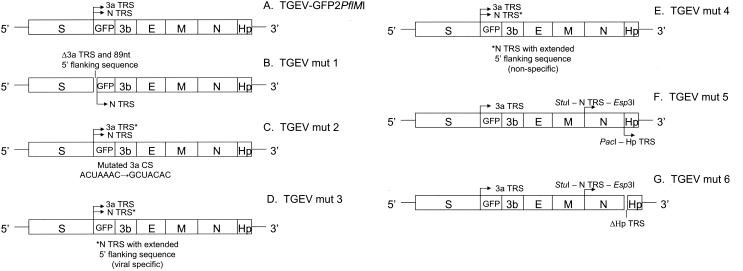
TGEV recombinant viruses expressing GFP. A series of TGEV recombinant viruses was generated using the TGEV-GFP2PflMI construct as the backbone for further characterization of the 3a and N gene TRS. (A) The TGEV-GFP2PflMI parent construct contains an ORF 3a deletion and insertion of GFP under the control of the 3a TRS and 20 nt of the N gene TRS. The location of the N and/or ORF 3a TRS sequences upstream of gfp is indicated. Clones were identified by DNA sequencing using an ABI model 377 automated sequencer, and the resulting constructs were subsequently used in the assembly of recombinant TGEV viral cDNAs. (B) TGEVmut1 was constructed by deleting nt 24709 to 24804 of the TGEV genome from TGEV-GFP2PflMI, corresponding to the ORF 3a CS and 89 nt of its 5′ flanking sequence. (C) TGEVmut2 contains the mutation of the upstream ORF 3a CS (ACUAAAC→GCUACAC), such that only the N CS is positioned upstream of gfp. (D) TGEVmut3 contains the 5′ N CS flanking sequence, which has been extended from 8 to 47 nt, (corresponding to nt 26858 to 26904 of the TGEV genome; GenBank accession no. AJ271965) while leaving the upstream ORF 3a CS completely intact. (E) TGEVmut4 contains the insertion of a nonspecific 39-nt sequence just 5′ of the 20 nt N gene TRS. (F) TGEVmut5 was engineered to contain unique StuI and Esp3I restriction sites which flank the CS element upstream of the N gene, and unique BstEII and PacI sites were introduced at the 3′ end of the N gene. The PacI restric-tion site was introduced at nucleotide position 28063 of the TGEV genome by mutagenesis of the Hp gene CS (ACTAAAC→ATTAATTAA). The wild-type Hp CS and gene were then repositioned just downstream of this PacI site, including 10 bp of upstream 5′ flanking sequence (nt 28051 to 28060). (G) To generate TGEVmut6, a ∼500-bp amplicon representing the CS deletion and 5′-truncated Hp gene was generated using the PacI site containing the 5′ primer (5′-TTA ATT AAA CCG GTT CGT CTT CCT CCA TGC TG-3′) and a 3′ primer within the cloning vector (Topo XL TA cloning vector; Invitrogen. Using the unique PacI site, this amplicon was inserted into the TGEVmut5 background to replace the wild-type Hp gene, resulting in a new recombinant TGEV F fragment containing a deletion of the CUAAAC Hp CS and 7 nt of 3′ flanking sequence (nt 28051 to 28074), including the first 4 nt of the Hp ORF. The only two available ATG start codons are out of frame with respect to the Hp ORF at nt positions 28087 and 28186 and would encode a seven- and nine-amino-acid protein, respectively.
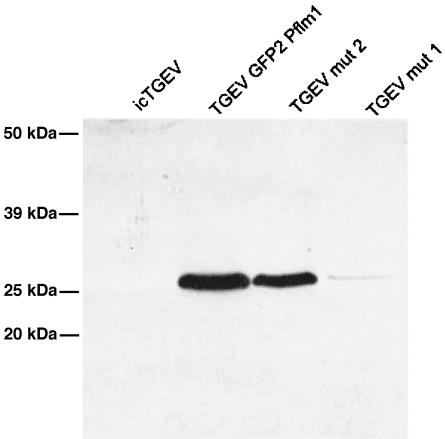
GFP expression from TGEV recombinant viruses. ST cells were infected with wild-type TGEV, TGEVmut2, or TGEVmut3, incubated for 12 h at 37°C, and subsequently lysed. Harvested lysates were subjected to Western blotting with monoclonal antisera directed against GFP (Clontech) or the TGEV N protein. Murine antisera against the TGEV N protein were raised by immunizing mice with Venezuelan equine encephalitis virus replicon vectors encoding the TGEV N gene.
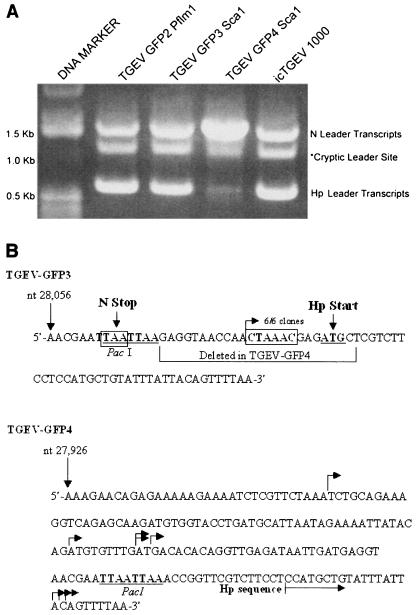
RT-PCR and sequence analysis of leader-containing transcripts in TGEV-GFP3- and TGEV-GFP4-infected cells. Cultures of ST cells were infected with TGEV-GFP2, -GFP3, or -GFP4 or wild-type TGEV derived from the infectious construct (icTGEV 1000) at a multiplicity of infection of sim for 1 h at room temperature. Intracellular RNA was harvested ∼12 h postinfection and used as a template for RT-PCR with the 5′ leader-specific primer TGEV-L (5′-CACTATTAGACTTTTAAAGTAAAGTGAGTGTAGC-3′) and a 3′ primer specific to the 3′ terminus of the TGEV genome (TGEV 3′ end; 5′-NNNNNNGCGGCCGCTTTTTTTTTTTTTTTTTTTTTTTTTGGTGTATCACTATCAAAAGG-3′). (A) Agarose gel electrophoresis of RT-PCR amplicons representing leader-containing subgenomic mRNA. RT-PCRs were run on 0.8% agarose gels, and amplicons of ∼1.5 kb and 600 bp were isolated, corresponding to leader-containing transcripts encoding N and Hp, respectively. A third amplicon of ∼1.3 kb was isolated that represents leader-containing transcripts driven from a cryptic start site. (B) Sequence analysis of RT-PCR amplicons from TGEV-GFP3- and TGEV-GFP4-infected cells. The ∼600 bp RT-PCR amplicons from TGEV-GFP3- and GFP4-infected cells shown in panel A were cloned and sequenced to determine the leader-body junctions. Leader-containing transcripts encoding Hp were exclusively initiated from the CS present just upstream of the Hp ORF in TGEV-GFP3 (six of six independent clones sequenced). Subgenomic mRNA synthesis was initiated from a variety of locations in TGEV-GFP4, ranging 124 nt upstream and 42 nt downstream from the original CS location in TGEV-GFP3 (eight independent clones sequence, each indicated by an arrow).
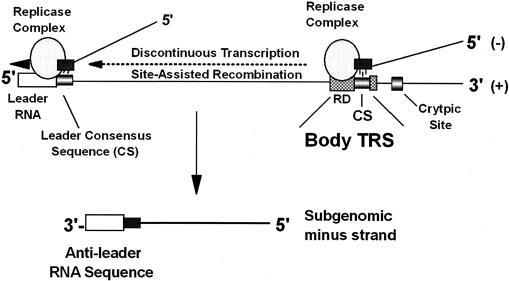
Model for the TGEV subgenomic TRS. The genomic RNA is the principle template for the synthesis of full-length and subgenomic-length negative-strand RNAs. Regulation of subgenomic negative-strand synthesis is mediated in part by proximity to the 3′ end of the genome and the presence of a TRS. The TRS is divided into RD and CS, both of which participate in discontinuous transcription. The regulatory domain of the TRS signals transcription attenuation, and the CS site allows for site-assisted recombination between complementary CS sequences located in incomplete nascent negative-strand RNAs and in the leader CS sequences encoded at the 5′ end of the genome. The nascent negative-strand RNAs are extended into complete negative strands by acquisition of anti-leader RNA sequences. In the absence of proximal body CS sites, nearby cryptic sites are inefficiently used in discontinuous transcription of subgenomic negative-strand RNAs.
Similar articles
-
Transcription regulatory sequences and mRNA expression levels in the coronavirus transmissible gastroenteritis virus.J Virol. 2002 Feb;76(3):1293-308. doi: 10.1128/jvi.76.3.1293-1308.2002. J Virol. 2002. PMID: 11773405 Free PMC article.
-
Role of nucleotides immediately flanking the transcription-regulating sequence core in coronavirus subgenomic mRNA synthesis.J Virol. 2005 Feb;79(4):2506-16. doi: 10.1128/JVI.79.4.2506-2516.2005. J Virol. 2005. PMID: 15681451 Free PMC article.
-
Sequence motifs involved in the regulation of discontinuous coronavirus subgenomic RNA synthesis.J Virol. 2004 Jan;78(2):980-94. doi: 10.1128/jvi.78.2.980-994.2004. J Virol. 2004. PMID: 14694129 Free PMC article.
-
Expression of transcriptional units using transmissible gastroenteritis coronavirus derived minigenomes and full-length cDNA clones.Adv Exp Med Biol. 2001;494:447-51. doi: 10.1007/978-1-4615-1325-4_65. Adv Exp Med Biol. 2001. PMID: 11774506 Review. No abstract available.
-
Coronavirus reverse genetics and development of vectors for gene expression.Curr Top Microbiol Immunol. 2005;287:161-97. doi: 10.1007/3-540-26765-4_6. Curr Top Microbiol Immunol. 2005. PMID: 15609512 Free PMC article. Review.
Cited by
-
Functional and genetic analysis of coronavirus replicase-transcriptase proteins.PLoS Pathog. 2005 Dec;1(4):e39. doi: 10.1371/journal.ppat.0010039. Epub 2005 Dec 9. PLoS Pathog. 2005. PMID: 16341254 Free PMC article. Review.
-
Coronaviruses: an RNA proofreading machine regulates replication fidelity and diversity.RNA Biol. 2011 Mar-Apr;8(2):270-9. doi: 10.4161/rna.8.2.15013. Epub 2011 Mar 1. RNA Biol. 2011. PMID: 21593585 Free PMC article. Review.
-
The molecular biology of coronaviruses.Adv Virus Res. 2006;66:193-292. doi: 10.1016/S0065-3527(06)66005-3. Adv Virus Res. 2006. PMID: 16877062 Free PMC article. Review.
-
Torovirus non-discontinuous transcription: mutational analysis of a subgenomic mRNA promoter.J Virol. 2005 Jul;79(13):8275-81. doi: 10.1128/JVI.79.13.8275-8281.2005. J Virol. 2005. PMID: 15956573 Free PMC article.
-
Reverse genetic manipulation of the overlapping coding regions for structural proteins of the type II porcine reproductive and respiratory syndrome virus.Virology. 2009 Jan 5;383(1):22-31. doi: 10.1016/j.virol.2008.09.013. Epub 2008 Nov 5. Virology. 2009. PMID: 18977502 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
