

Preferential binding to branched DNA strands and strand-annealing activity of the human Rad51B, Rad51C, Rad51D and Xrcc2 protein complex
- PMID: 15141025
- PMCID: PMC419466
- DOI: 10.1093/nar/gkh578
Preferential binding to branched DNA strands and strand-annealing activity of the human Rad51B, Rad51C, Rad51D and Xrcc2 protein complex
Abstract
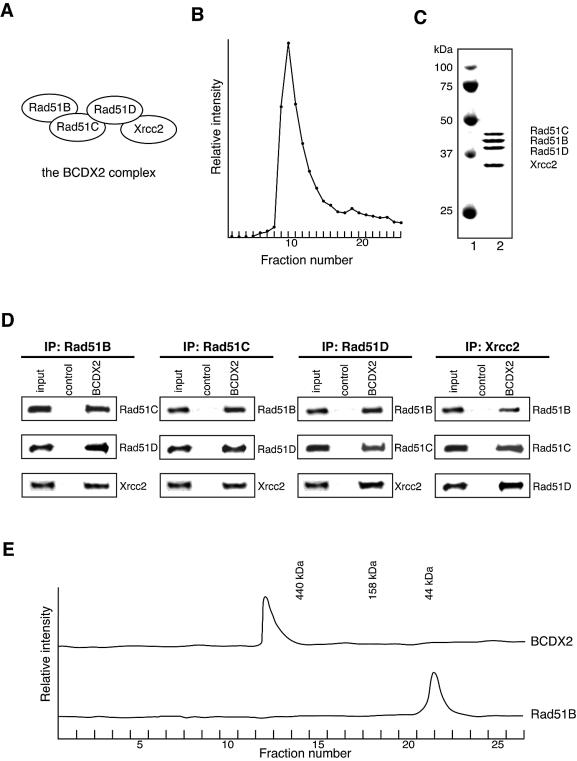
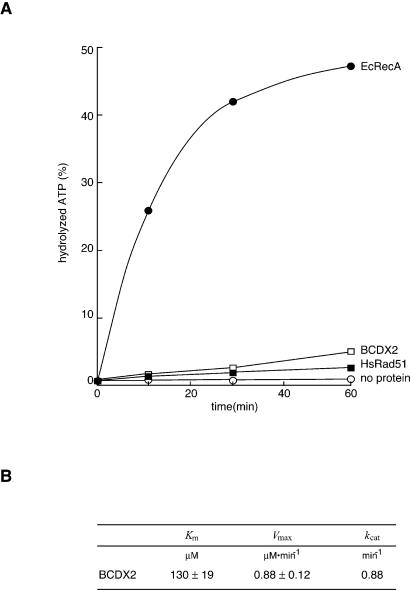
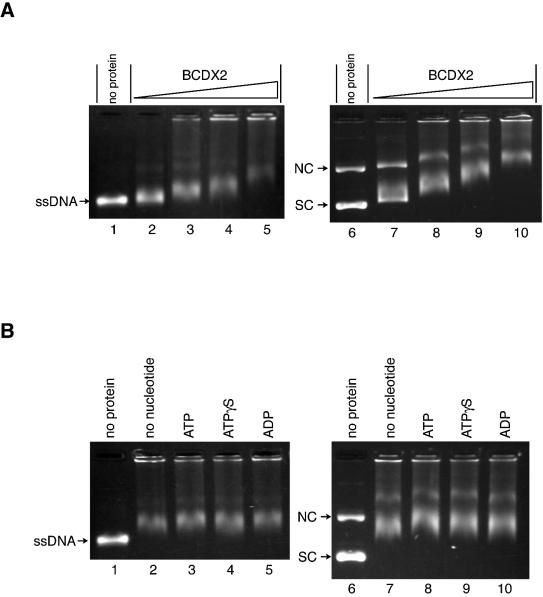
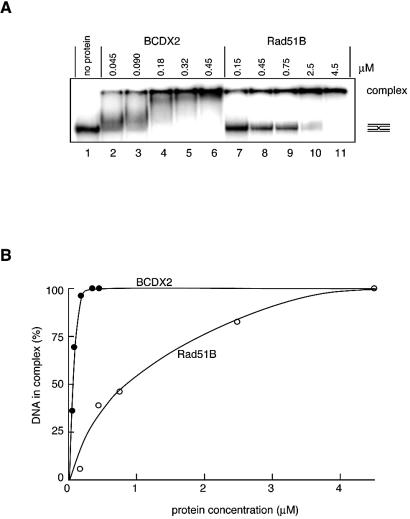
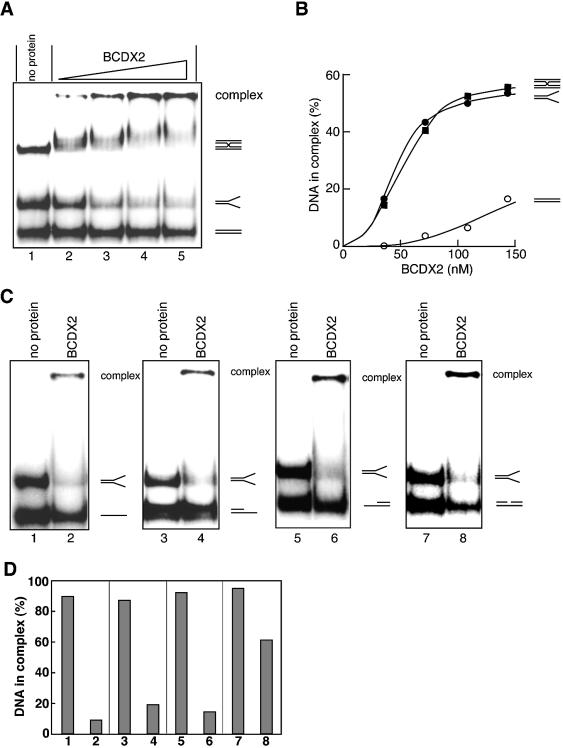
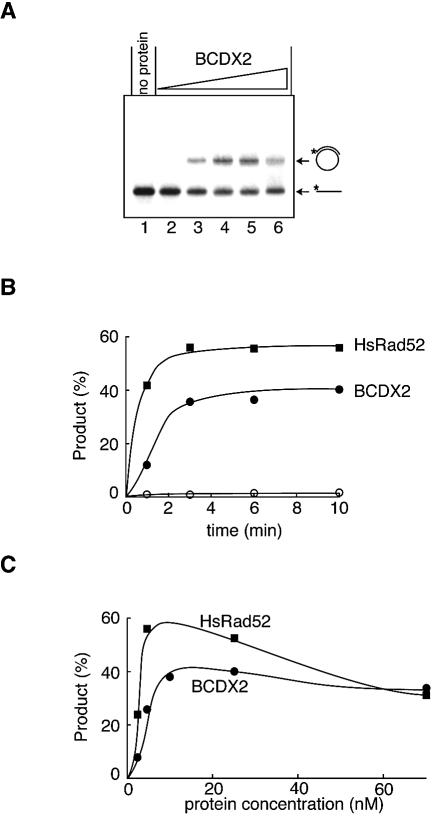
The Rad51B, Rad51C, Rad51D and Xrcc2 proteins are Rad51 paralogs, and form a complex (BCDX2 complex) in mammalian cells. Mutant cells defective in any one of the Rad51-paralog genes exhibit spontaneous genomic instability and extreme sensitivity to DNA-damaging agents, due to inefficient recombinational repair. Therefore, the Rad51 paralogs play important roles in the maintenance of genomic integrity through recombinational repair. In the present study, we examined the DNA-binding preference of the human BCDX2 complex. Competitive DNA-binding assays using seven types of DNA substrates, single-stranded DNA (ssDNA), double-stranded DNA, 5'- and 3'-tailed duplexes, nicked duplex DNA, Y-shaped DNA and a synthetic Holliday junction, revealed that the BCDX2 complex preferentially bound to the two DNA substrates with branched structures (the Y-shaped DNA and the synthetic Holliday junction). Furthermore, the BCDX2 complex catalyzed the strand-annealing reaction between a long linear ssDNA (1.2 kb in length) and its complementary circular ssDNA. These properties of the BCDX2 complex may be important for its roles in the maintenance of chromosomal integrity.
Figures
References
-
- Whitaker S.J. (1992) DNA damage by drugs and radiation: what is important and how is it measured? Eur. J. Cancer, 28, 273–276. - PubMed
-
- Cox M.M., Goodman,M.F., Kreuzer,K.N., Sherratt,D.J., Sandler,S.J. and Marians,K.J. (2000) The importance of repairing stalled replication forks. Nature, 404, 37–41. - PubMed
-
- Ward J.F. (1994) The complexity of DNA damage: relevance to biological consequences. Int. J. Radiat. Biol., 66, 427–432. - PubMed
-
- Caldecott K.W. (2001) Mammalian DNA single-strand break repair: an X-ra(y)ted affair. Bioessays, 23, 447–455. - PubMed
-
- van den Bosch M., Lohman,P.H. and Pastink,A. (2002) DNA double-strand break repair by homologous recombination. Biol. Chem., 383, 873–892. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
