

Synapsis and DNA cleavage in phiC31 integrase-mediated site-specific recombination
- PMID: 15141031
- PMCID: PMC419440
- DOI: 10.1093/nar/gkh538
Synapsis and DNA cleavage in phiC31 integrase-mediated site-specific recombination
Abstract
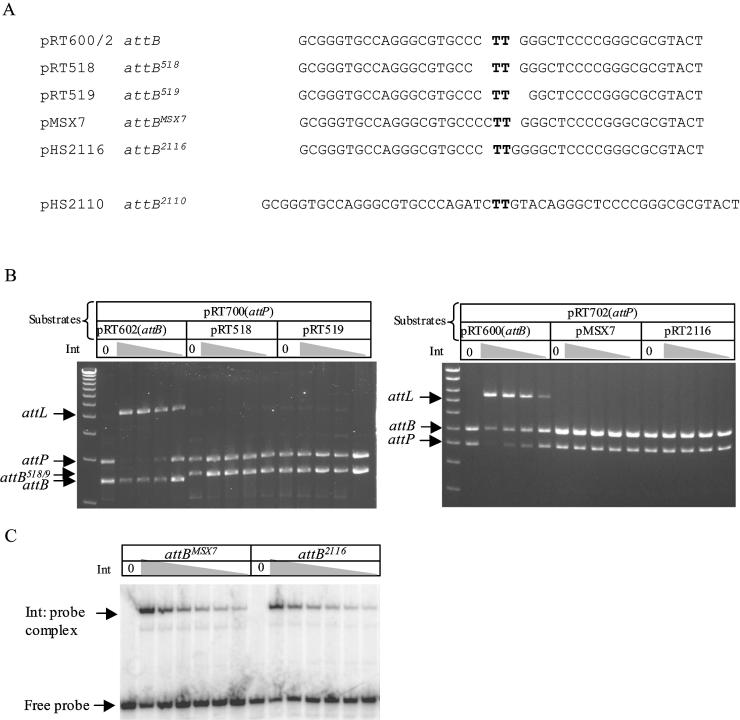
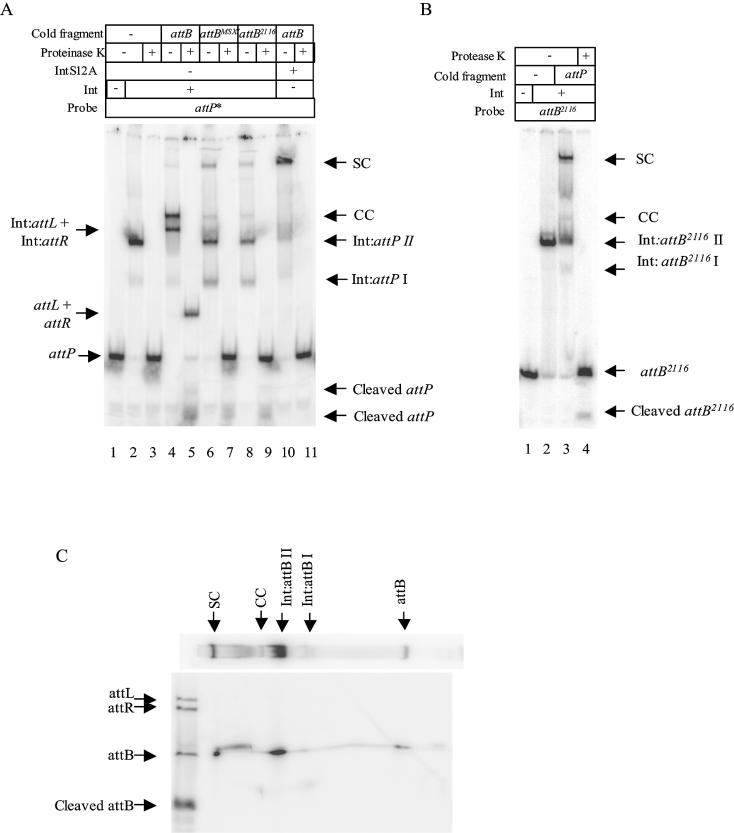
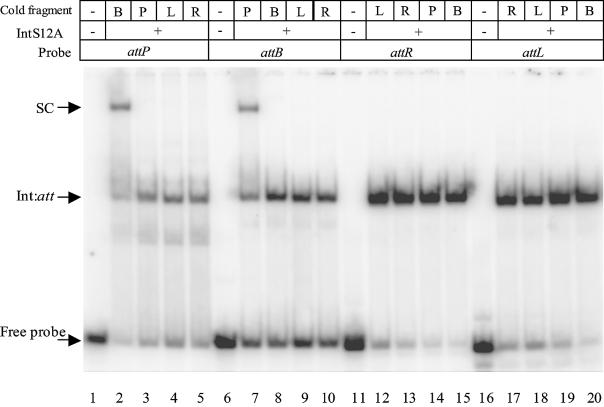
The Streptomyces phage phiC31 encodes an integrase belonging to the serine recombinase family of site-specific recombinases. The well studied serine recombinases, the resolvase/invertases, bring two recombination sites together in a synapse, and then catalyse a concerted four-strand staggered break in the DNA substrates whilst forming transient covalent attachments with the recessed 5' ends. Rotation of one pair of half sites by 180 degrees relative to the other pair occurs, to form the recombinant configuration followed by ligation of the DNA backbone. Here we address the nature of the recombination intermediates formed by phiC31 integrase when acting on its substrates attP and attB. We have identified intermediates containing integrase covalently attached to cleaved DNA substrates, attB or attP, by analysis of complexes in gels and after treatment of these complexes with proteinases. Using a catalytically inactive integrase mutant, S12A, the synaptic complexes containing integrase, attP and attB were identified. Furthermore, we have shown that attB mutants containing insertions or deletions are blocked in recombination at the stage of strand cleavage. Thus, there is a strict spacing requirement within attB, possibly for correct positioning of the catalytic serine relative to the scissile phosphate in the active site. Finally, using integrase S12A we confirmed the inability of attL and attR or other combinations of sites to form a stable synapse, indicating that the directionality of integrative recombination is determined at synapsis.
Figures
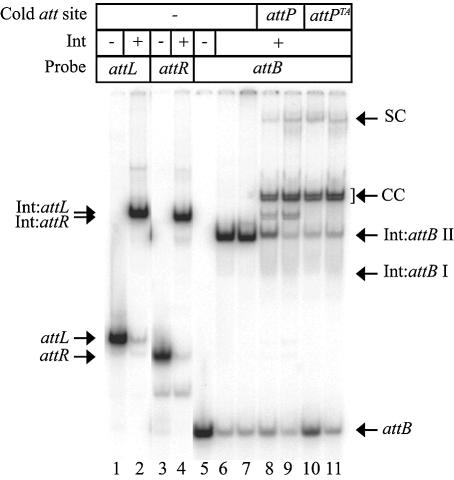
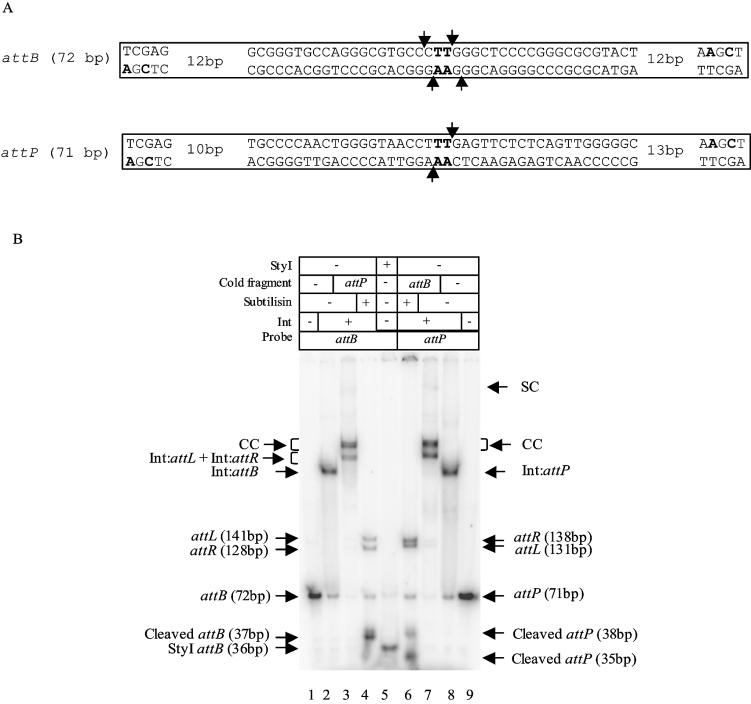
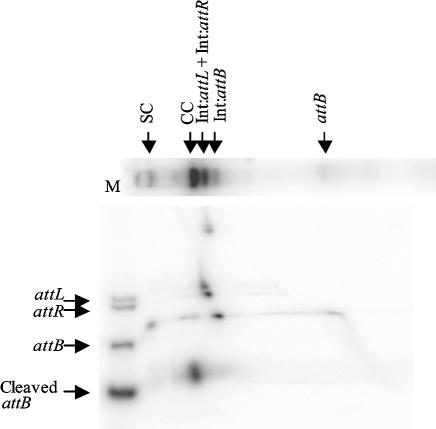
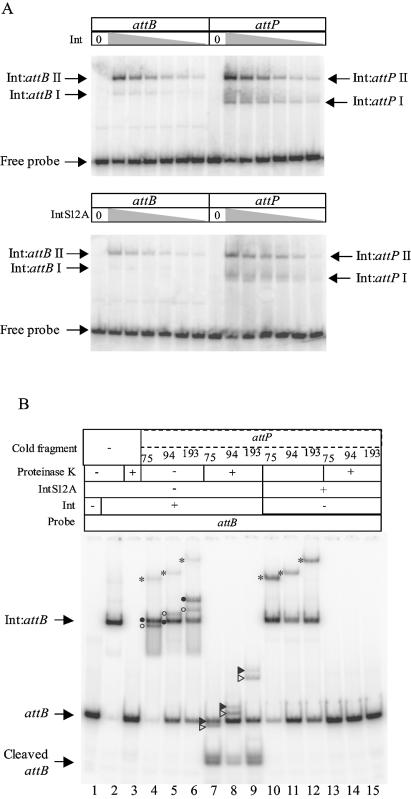
References
-
- Smith M.C.M. and Thorpe,H.M. (2002) Diversity in the serine recombinases. Mol. Microbiol., 44, 299–307. - PubMed
-
- Kunkel B., Losick,R. and Stragier,P. (1990) The Bacillus subtilis gene for the development transcription factor sigma K is generated by excision of a dispensable DNA element containing a sporulation recombinase gene. Genes Dev., 4, 525–535. - PubMed
-
- Carrasco C.D., Ramaswamy,K.S., Ramasubramanian,T.S. and Golden,J.W. (1994) Anabaena xisF gene encodes a developmentally regulated site-specific recombinase. Genes Dev., 8, 74–83. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
