

Neuroactive steroids have multiple actions to potentiate GABAA receptors
- PMID: 15146041
- PMCID: PMC1664905
- DOI: 10.1113/jphysiol.2004.066571
Neuroactive steroids have multiple actions to potentiate GABAA receptors
Abstract
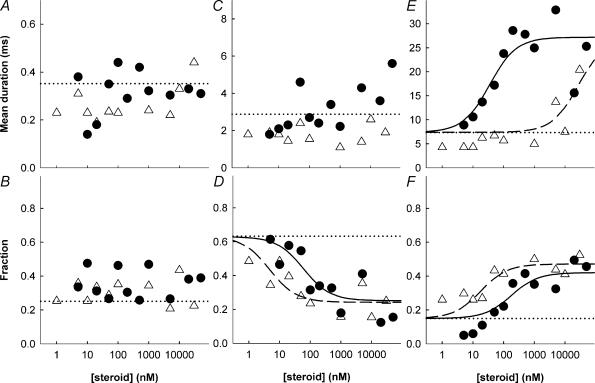
The effects of neuroactive steroids on the function of GABAA receptors were studied using cell-attached records of single channel activity recorded from HEK293 cells transfected with alpha1 beta2 gamma2L subunits. Activity was elicited with a half-maximal (50 microM) concentration of GABA. Two steroids were studied in detail: ACN ((3alpha,5alpha,17beta)-3-hydroxyandrostane-17-carbonitrile) and B285 ((3alpha,5beta,17beta)-3-hydroxy-18-norandrostane-17-carbonitrile). Four effects on channel activity were seen, two on open time distributions and two on closed times. When clusters of openings were elicited in the absence of steroid, the open time distribution contained three components. ACN produced concentration-dependent alterations in the open time distribution. The prevalence of the longest duration class of open times was increased from about 15% to about 40% (EC50 about 180 nM ACN), while the duration of the longest class increased from 7.4 ms to 27 ms (EC50 about 35 nM ACN). B285 also increased the prevalence of the longest duration open times (EC50 about 18 nM B285) but increased the duration only at concentrations close to 10 microM. The differences in the actions of these two steroids suggest that the effects on proportion and duration of the long duration open time component are produced by independent mechanisms and that there are separate recognition sites for the steroids which are associated with the two functional actions. The closed time distributions also showed three components in the absence of steroid. The rate of occurrence of the two brief duration closed time components decreased with increasing ACN, with an EC50 of about 50 nM ACN. In contrast, B285 did not reduce the rate of occurrence of the brief closings until high concentrations were applied. However, both B285 and ACN reduced the rate of occurrence of the activation-related closed state selectively, with comparable IC50 concentrations (about 40 nM ACN, 20 nM B285). As in the case for action on open times these data suggest that there are two recognition sites and two independent mechanisms, perhaps the sites and mechanisms associated with actions on open times. The presence of 1 microM ACN had no effect on the estimated channel opening rate or on the apparent affinity of the receptor for GABA. Mutation of the carboxy terminus of the gamma2 subunit, but not the alpha1 or beta2 subunits, abolished the ability of ACN to increase the duration of OT3 but had no effect on the reduction of the rate of occurrence of the activation-related closed state. These observations are also consistent with the idea that there is more than one distinguishable steroid recognition site on the GABAA receptor.
Figures








References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials