

Diabetes and exocrine pancreatic insufficiency in E2F1/E2F2 double-mutant mice
- PMID: 15146237
- PMCID: PMC406522
- DOI: 10.1172/JCI18879
Diabetes and exocrine pancreatic insufficiency in E2F1/E2F2 double-mutant mice
Abstract
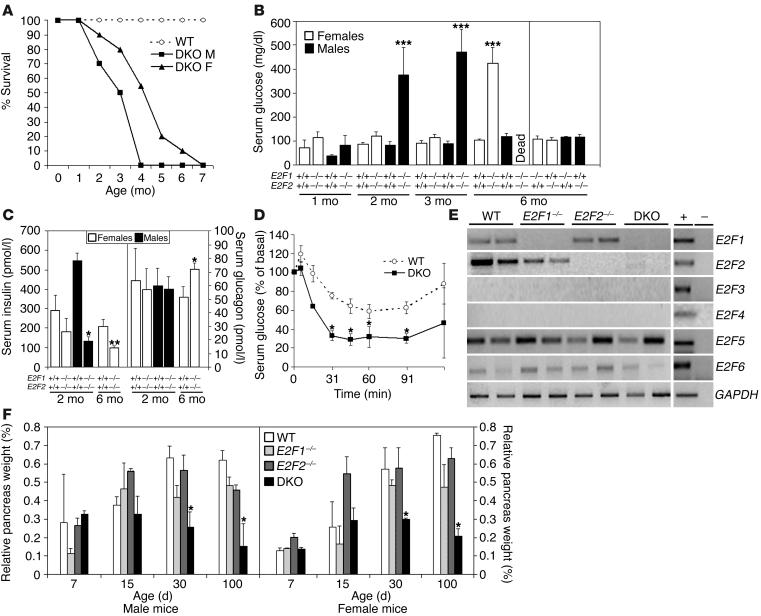
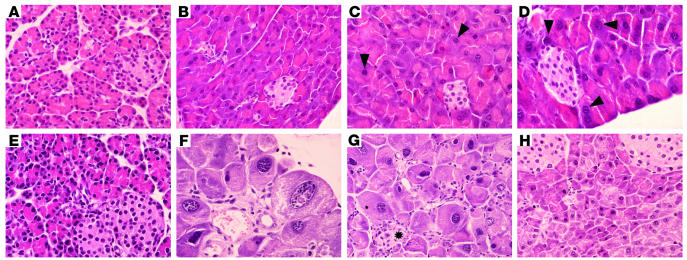
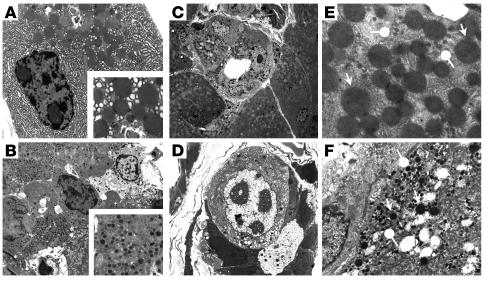
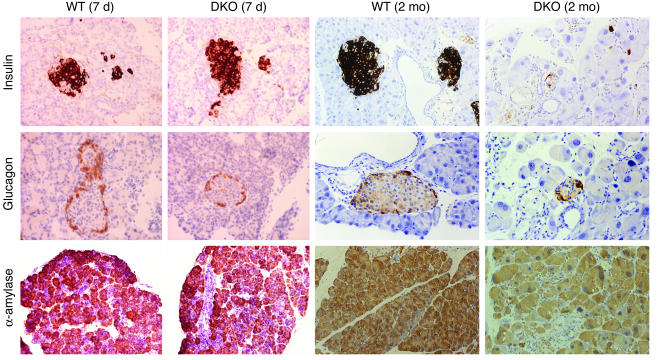
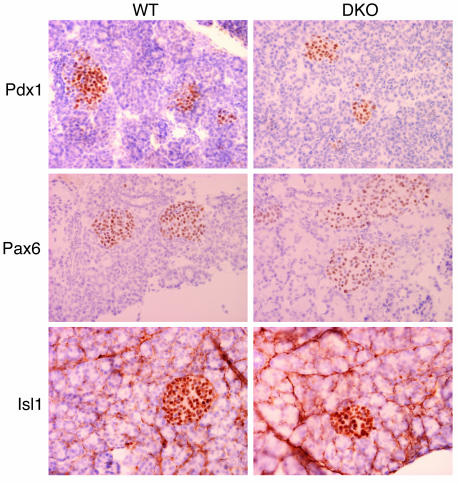
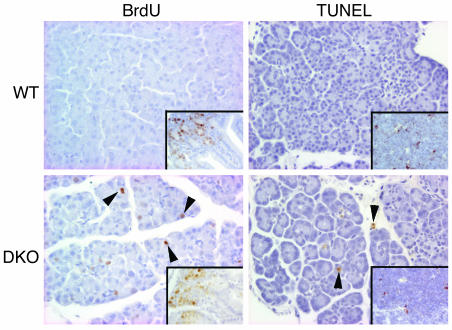
E2F transcription factors are thought to be key regulators of cell growth control. Here we use mutant mouse strains to investigate the function of E2F1 and E2F2 in vivo. E2F1/E2F2 compound-mutant mice develop nonautoimmune insulin-deficient diabetes and exocrine pancreatic dysfunction characterized by endocrine and exocrine cell dysplasia, a reduction in the number and size of acini and islets, and their replacement by ductal structures and adipose tissue. Mutant pancreatic cells exhibit increased rates of DNA replication but also of apoptosis, resulting in severe pancreatic atrophy. The expression of genes involved in DNA replication and cell cycle control was upregulated in the E2F1/E2F2 compound-mutant pancreas, suggesting that their expression is repressed by E2F1/E2F2 activities and that the inappropriate cell cycle found in the mutant pancreas is likely the result of the deregulated expression of these genes. Interestingly, the expression of ductal cell and adipocyte differentiation marker genes was also upregulated, whereas expression of pancreatic cell marker genes were downregulated. These results suggest that E2F1/E2F2 activity negatively controls growth of mature pancreatic cells and is necessary for the maintenance of differentiated pancreatic phenotypes in the adult.
Figures
Similar articles
-
Activation of ARF by oncogenic stress in mouse fibroblasts is independent of E2F1 and E2F2.Oncogene. 2002 May 2;21(19):2939-47. doi: 10.1038/sj.onc.1205371. Oncogene. 2002. PMID: 12082524
-
Specificity of E2F1, E2F2, and E2F3 in mediating phenotypes induced by loss of Rb.Cell Growth Differ. 2002 May;13(5):215-25. Cell Growth Differ. 2002. PMID: 12065245
-
Unique roles for E2F1 in the mouse lens in the absence of functional pRB proteins.Invest Ophthalmol Vis Sci. 2002 May;43(5):1509-16. Invest Ophthalmol Vis Sci. 2002. PMID: 11980867
-
E2F1 in gliomas: a paradigm of oncogene addiction.Cancer Lett. 2008 May 18;263(2):157-63. doi: 10.1016/j.canlet.2008.02.001. Epub 2008 Mar 10. Cancer Lett. 2008. PMID: 18334281 Review.
-
[Knockout mouse in the research of type 1 diabetes].Nihon Rinsho. 2002 Aug;60 Suppl 8:52-7. Nihon Rinsho. 2002. PMID: 12355801 Review. Japanese. No abstract available.
Cited by
-
Survey of the human pancreatic beta-cell G1/S proteome reveals a potential therapeutic role for cdk-6 and cyclin D1 in enhancing human beta-cell replication and function in vivo.Diabetes. 2009 Apr;58(4):882-93. doi: 10.2337/db08-0631. Epub 2009 Jan 9. Diabetes. 2009. PMID: 19136653 Free PMC article.
-
Ablation of the transcription factors E2F1-2 limits neuroinflammation and associated neurological deficits after contusive spinal cord injury.Cell Cycle. 2015;14(23):3698-712. doi: 10.1080/15384101.2015.1104436. Cell Cycle. 2015. PMID: 26505089 Free PMC article.
-
E2F1 and E2F2 prevent replicative stress and subsequent p53-dependent organ involution.Cell Death Differ. 2015 Oct;22(10):1577-89. doi: 10.1038/cdd.2015.4. Epub 2015 Feb 6. Cell Death Differ. 2015. PMID: 25656653 Free PMC article.
-
Rb and p107 are required for alpha cell survival, beta cell cycle control and glucagon-like peptide-1 action.Diabetologia. 2014 Dec;57(12):2555-65. doi: 10.1007/s00125-014-3381-y. Epub 2014 Sep 24. Diabetologia. 2014. PMID: 25249236
-
The pRb/RBL2-E2F1/4-GCN5 axis regulates cancer stem cell formation and G0 phase entry/exit by paracrine mechanisms.Nat Commun. 2024 Apr 27;15(1):3580. doi: 10.1038/s41467-024-47680-z. Nat Commun. 2024. PMID: 38678032 Free PMC article.
References
-
- Boyd KE, Farnham PJ. Identification of target genes of oncogenic transcription factors. Proc. Soc. Exp. Biol. Med. 1999;222:9–28. - PubMed
-
- Dyson N. The regulation of E2F by pRb-family proteins. Genes Dev. 1998;12:2245–2262. - PubMed
-
- Helin K. Regulation of cell proliferation by the E2F transcription factors. Curr. Opin. Genet. Dev. 1998;8:28–35. - PubMed
-
- DeGregori J. The genetics of the E2F family of transcription factors: shared functions and unique roles. . Biochim. Biophys. Acta. 2002; 1602:131–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
