

CD8 T cell recognition of endogenously expressed epstein-barr virus nuclear antigen 1
- PMID: 15148339
- PMCID: PMC2211813
- DOI: 10.1084/jem.20040121
CD8 T cell recognition of endogenously expressed epstein-barr virus nuclear antigen 1
Abstract
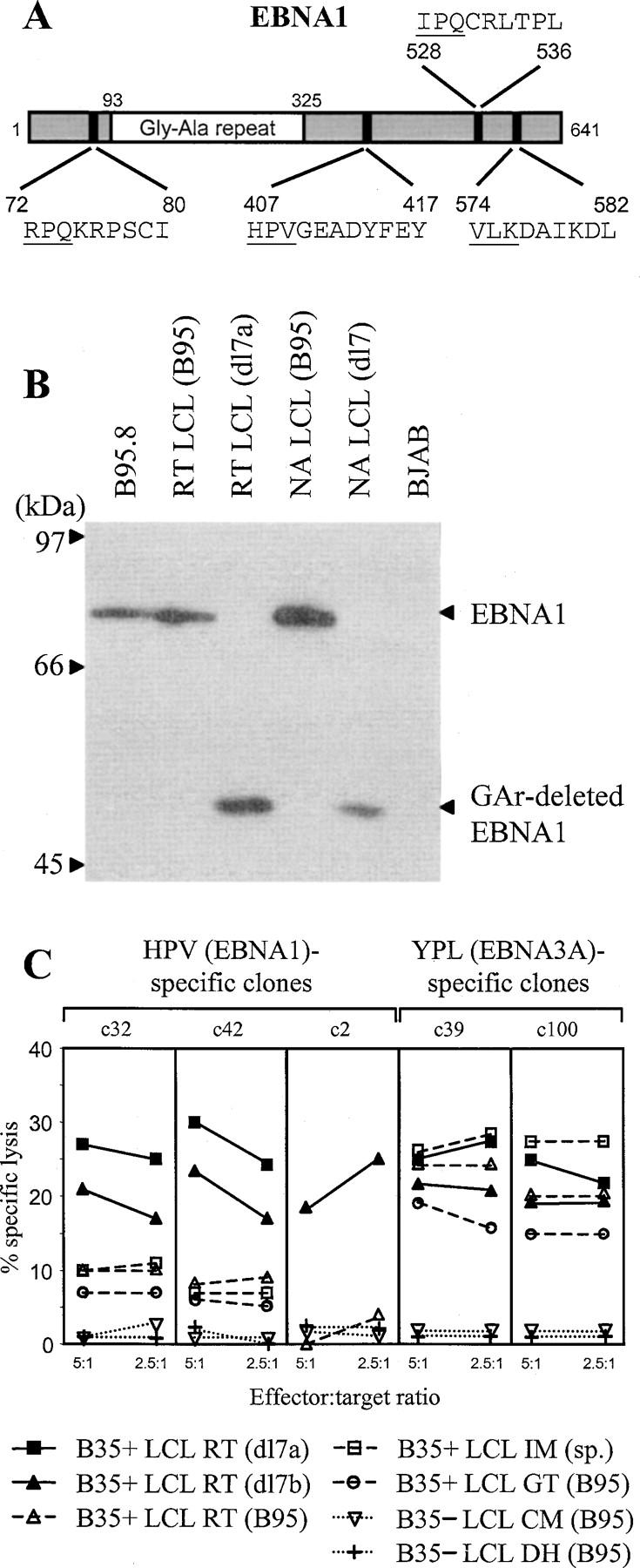
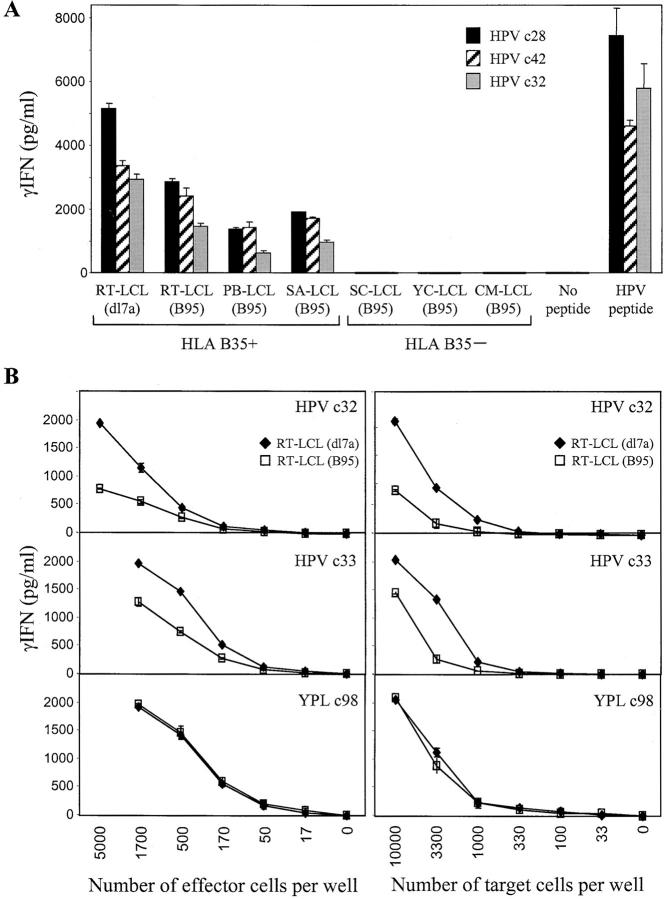
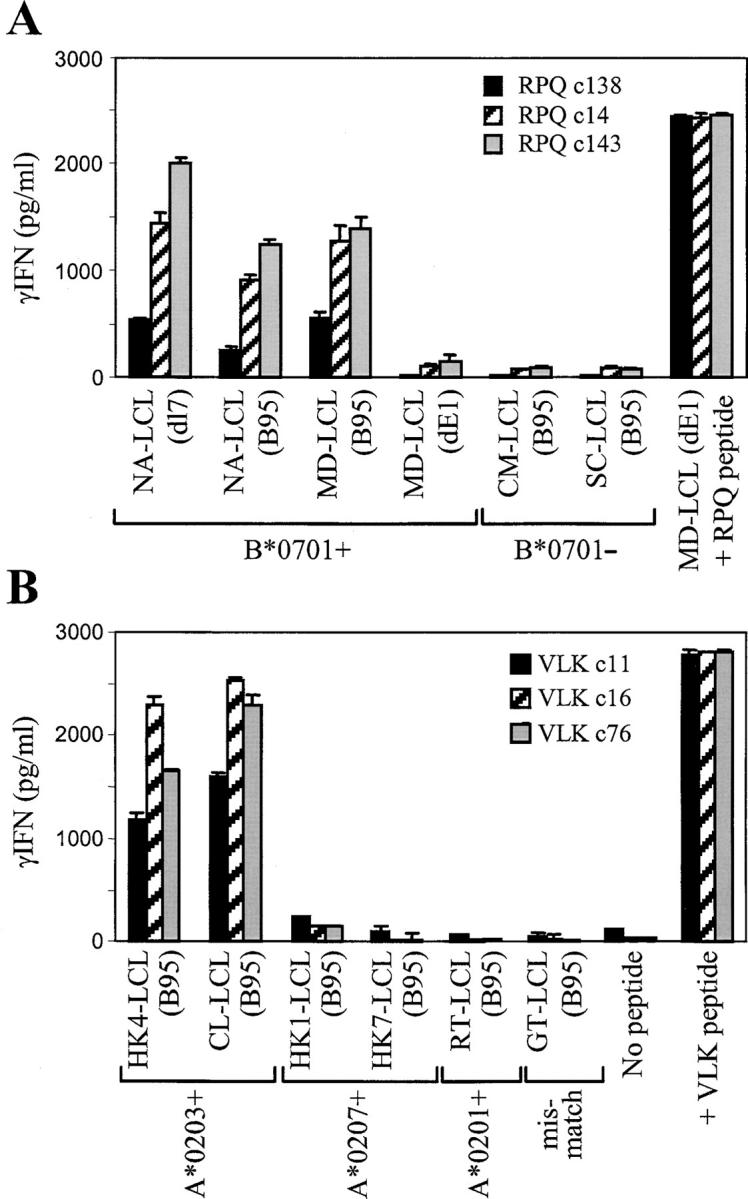
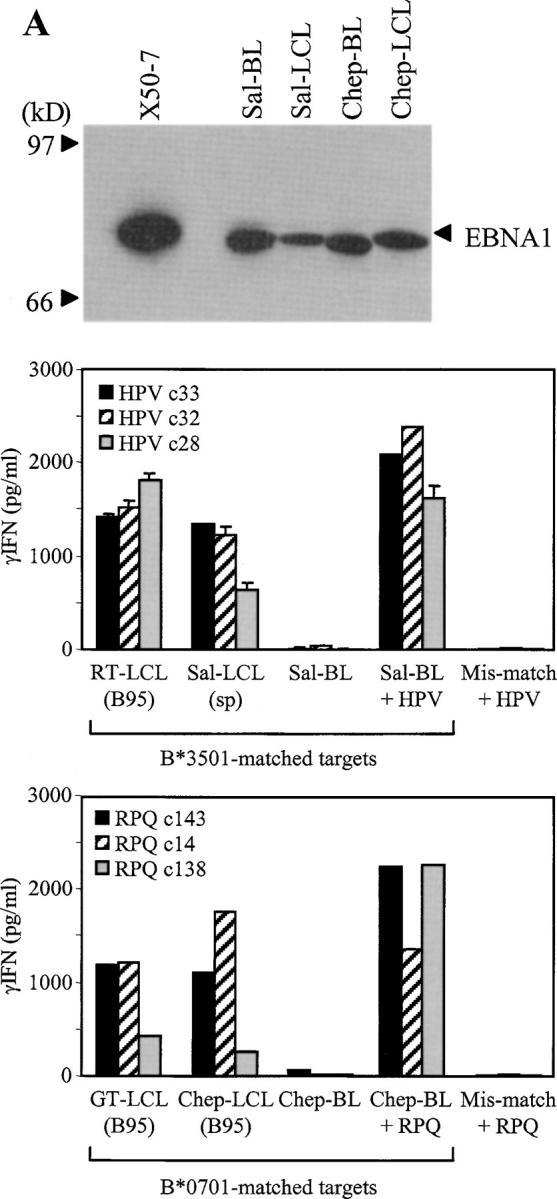
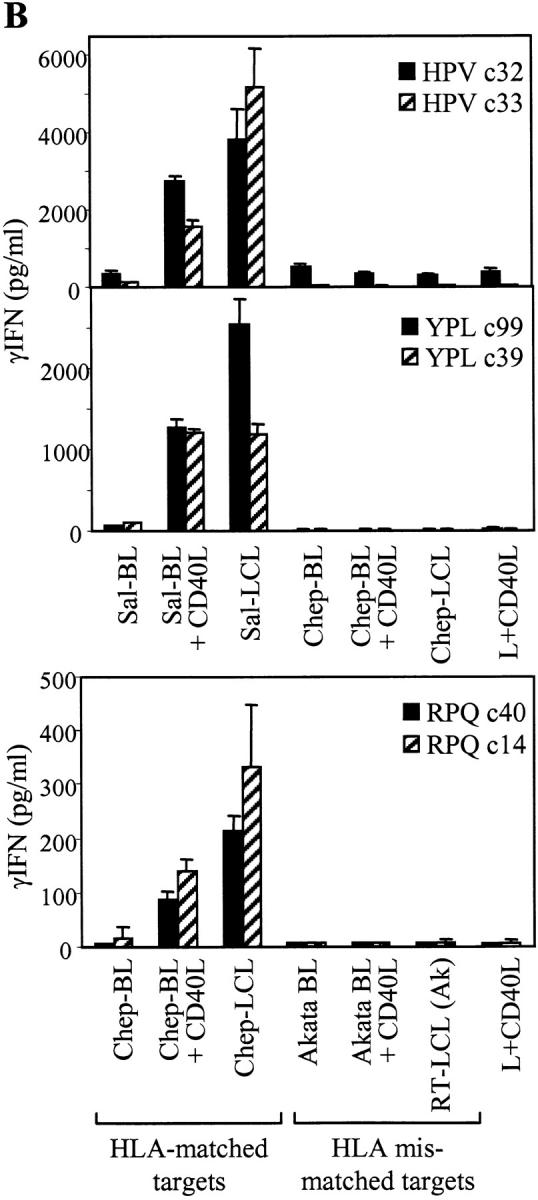
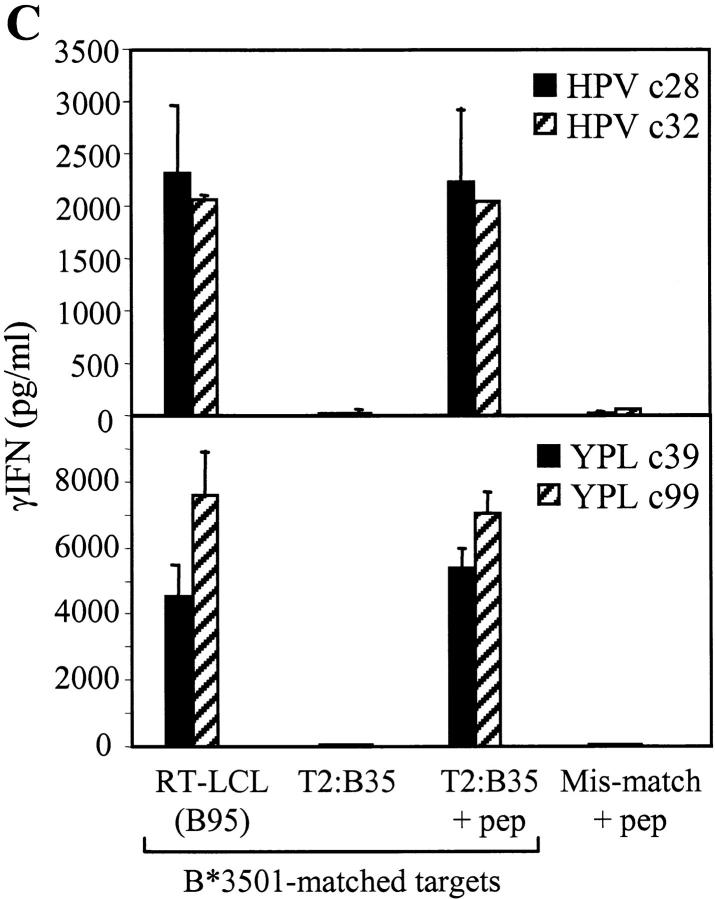
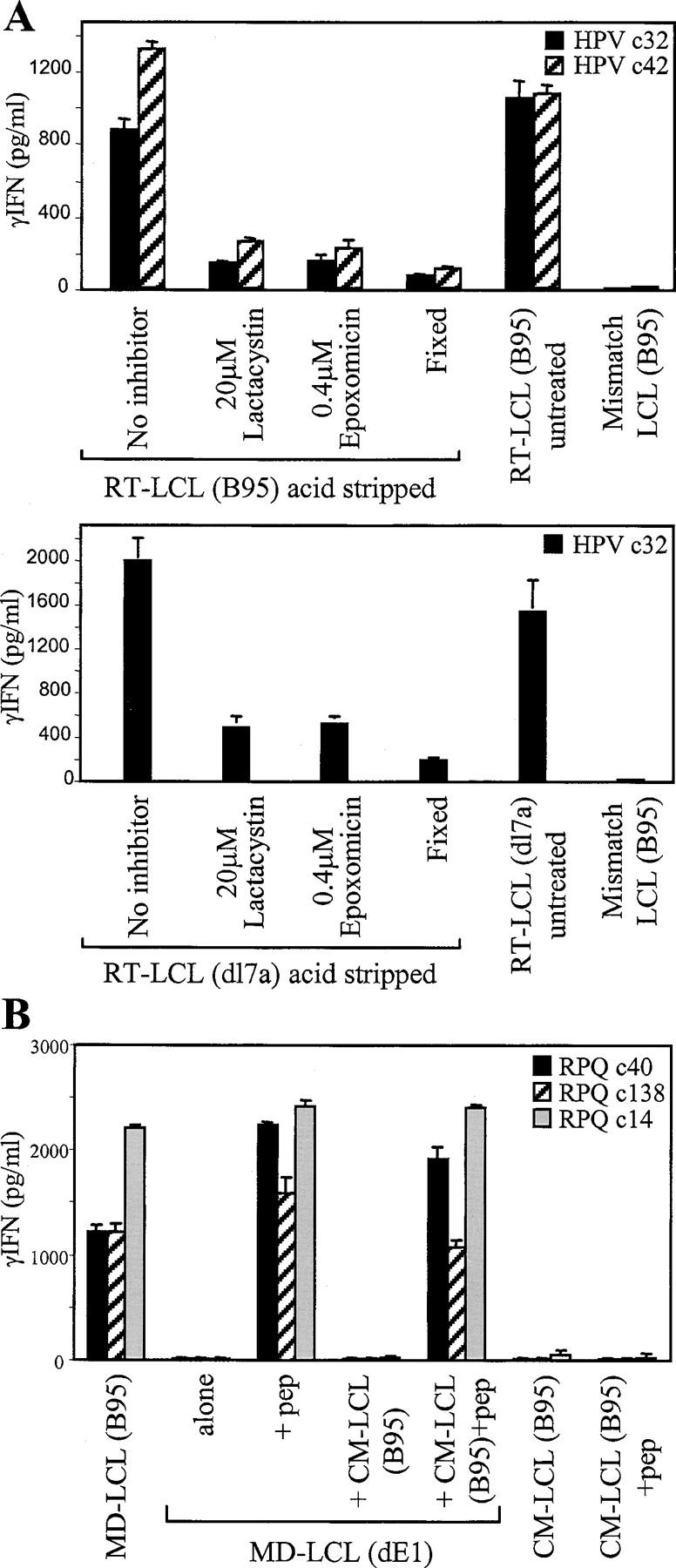
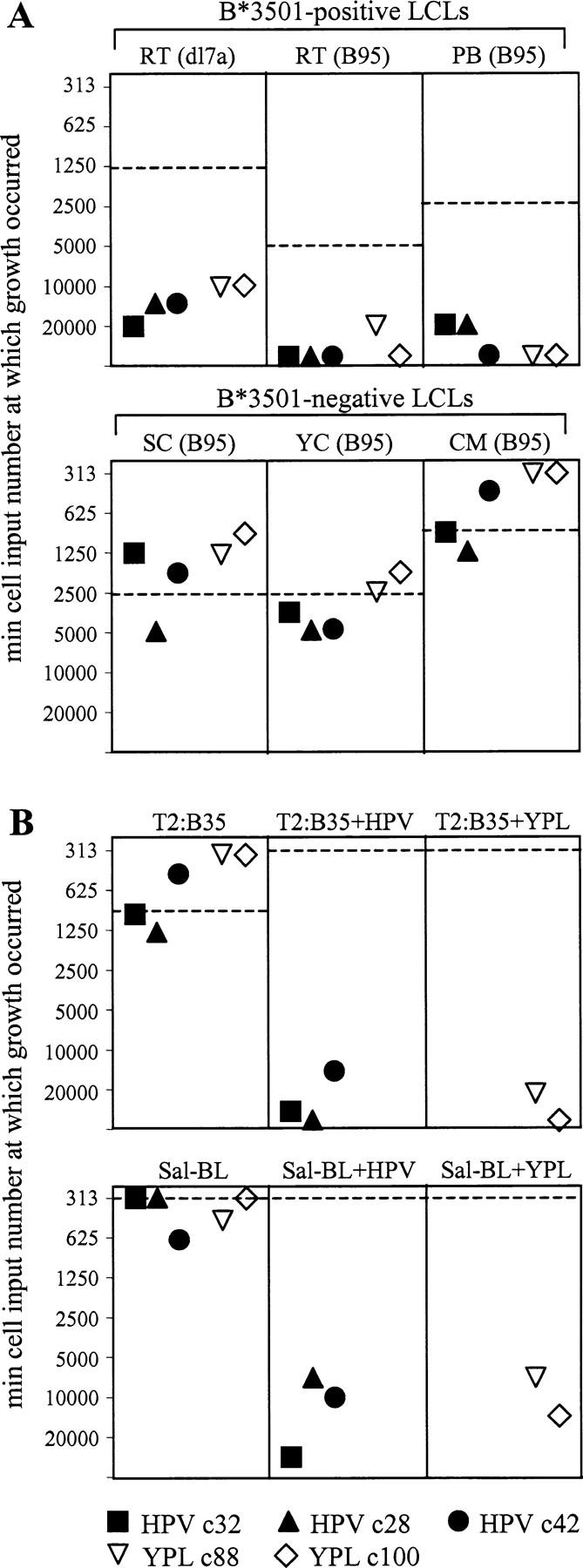
The Epstein-Barr virus (EBV) nuclear antigen (EBNA)1 contains a glycine-alanine repeat (GAr) domain that appears to protect the antigen from proteasomal breakdown and, as measured in cytotoxicity assays, from major histocompatibility complex (MHC) class I-restricted presentation to CD8+ T cells. This led to the concept of EBNA1 as an immunologically silent protein that although unique in being expressed in all EBV malignancies, could not be exploited as a CD8 target. Here, using CD8+ T cell clones to native EBNA1 epitopes upstream and downstream of the GAr domain and assaying recognition by interferon gamma release, we show that the EBNA1 naturally expressed in EBV-transformed lymphoblastoid cell lines (LCLs) is in fact presented to CD8+ T cells via a proteasome/peptide transporter-dependent pathway. Furthermore, LCL recognition by such CD8+ T cells, although slightly lower than seen with paired lines expressing a GAr-deleted EBNA1 protein, leads to strong and specific inhibition of LCL outgrowth in vitro. Endogenously expressed EBNA1 is therefore accessible to the MHC class I pathway despite GAr-mediated stabilization of the mature protein. We infer that EBNA1-specific CD8+ T cells do play a role in control of EBV infection in vivo and might be exploitable in the control of EBV+ malignancies.
Figures
References
-
- Pamer, E., and P. Cresswell. 1998. Mechanisms of MHC class I–restricted antigen processing. Annu. Rev. Immunol. 16:323–358. - PubMed
-
- Koszinowski, U.H., and H. Hengel. 2002. Viral Proteins Counteracting Host Defences. Springer-Verlag, Berlin. 325 pp.
-
- Rickinson, A.B., and E. Kieff. 2001. Epstein-Barr virus. Fields Virology. D.M. Knipe, and P.M. Howley, editors. Lippincott Williams and Wilkins, Philadelphia. 2575–2627.
-
- Levitskaya, J., M. Coram, V. Levitsky, S. Imreh, P.M. Steigerwald-Mullen, G. Klein, M.G. Kurilla, and M.G. Masucci. 1995. Inhibition of antigen processing by the internal repeat region of the Epstein-Barr virus nuclear antigen 1. Nature. 375:685–688. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
