

Transcription by an archaeal RNA polymerase is slowed but not blocked by an archaeal nucleosome
- PMID: 15150236
- PMCID: PMC415759
- DOI: 10.1128/JB.186.11.3492-3498.2004
Transcription by an archaeal RNA polymerase is slowed but not blocked by an archaeal nucleosome
Abstract
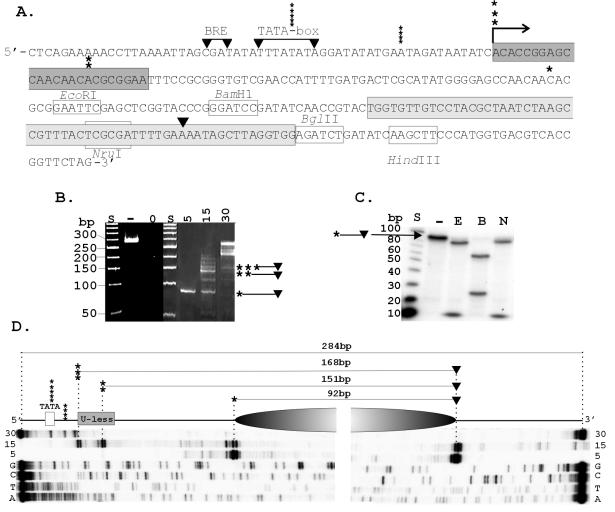
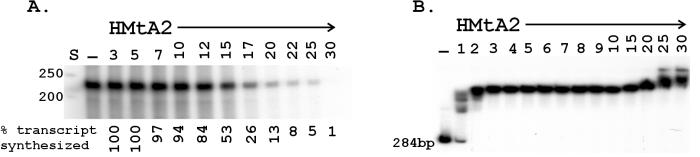
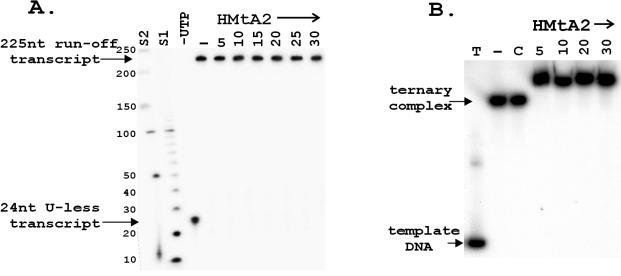
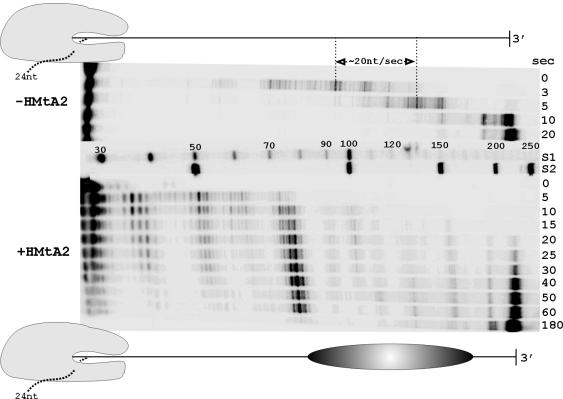
Archaeal RNA polymerases (RNAPs) are closely related to eukaryotic RNAPs, and in Euryarchaea, genomic DNA is wrapped and compacted by histones into archaeal nucleosomes. In eukaryotes, transcription of DNA bound into nucleosomes is facilitated by histone tail modifications and chromatin remodeling complexes, but archaeal histones do not have histone tails and archaeal genome sequences provide no evidence for archaeal homologs of eukaryotic chromatin remodeling complexes. We have therefore investigated the ability of an archaeal RNAP, purified from Methanothermobacter thermautotrophicus, to transcribe DNA bound into an archaeal nucleosome by HMtA2, an archaeal histone from M. thermautotrophicus. To do so, we constructed a template that allows transcript elongation to be separated from transcription initiation, on which archaeal nucleosome assembly is positioned downstream from the site of transcription initiation. At 58 degrees C, in the absence of an archaeal nucleosome, M. thermautotrophicus RNAP transcribed this template DNA at a rate of approximately 20 nucleotides per second. With an archaeal nucleosome present, transcript elongation was slowed but not blocked, with transcription pausing at sites before and within the archaeal nucleosome. With additional HMtA2 binding, complexes were obtained that also incorporated the upstream regulatory region. This inhibited transcription presumably by preventing archaeal TATA-box binding protein, general transcription factor TFB, and RNAP access and thus inhibiting transcription initiation.
Figures
Similar articles
-
Transcription by Methanothermobacter thermautotrophicus RNA polymerase in vitro releases archaeal transcription factor B but not TATA-box binding protein from the template DNA.J Bacteriol. 2004 Sep;186(18):6306-10. doi: 10.1128/JB.186.18.6306-6310.2004. J Bacteriol. 2004. PMID: 15342601 Free PMC article.
-
Archaeal RNA polymerase is sensitive to intrinsic termination directed by transcribed and remote sequences.J Mol Biol. 2006 Jan 13;355(2):196-210. doi: 10.1016/j.jmb.2005.10.062. Epub 2005 Nov 9. J Mol Biol. 2006. PMID: 16305799
-
Archaeal nucleosome positioning in vivo and in vitro is directed by primary sequence motifs.BMC Genomics. 2013 Jun 10;14:391. doi: 10.1186/1471-2164-14-391. BMC Genomics. 2013. PMID: 23758892 Free PMC article.
-
Archaeal chromatin and transcription.Mol Microbiol. 2003 May;48(3):587-98. doi: 10.1046/j.1365-2958.2003.03439.x. Mol Microbiol. 2003. PMID: 12694606 Review.
-
Changing the DNA landscape: putting a SPN on chromatin.Curr Top Microbiol Immunol. 2003;274:171-201. doi: 10.1007/978-3-642-55747-7_7. Curr Top Microbiol Immunol. 2003. PMID: 12596908 Review.
Cited by
-
DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius.Biomolecules. 2022 Mar 30;12(4):524. doi: 10.3390/biom12040524. Biomolecules. 2022. PMID: 35454113 Free PMC article.
-
Regulation of tryptophan operon expression in the archaeon Methanothermobacter thermautotrophicus.J Bacteriol. 2005 Sep;187(18):6419-29. doi: 10.1128/JB.187.18.6419-6429.2005. J Bacteriol. 2005. PMID: 16159776 Free PMC article.
-
Chromatinization of Escherichia coli with archaeal histones.Elife. 2019 Nov 6;8:e49038. doi: 10.7554/eLife.49038. Elife. 2019. PMID: 31692448 Free PMC article.
-
The interplay between nucleoid organization and transcription in archaeal genomes.Nat Rev Microbiol. 2015 Jun;13(6):333-41. doi: 10.1038/nrmicro3467. Epub 2015 May 6. Nat Rev Microbiol. 2015. PMID: 25944489 Review.
-
The DNA-binding protein HTa from Thermoplasma acidophilum is an archaeal histone analog.Elife. 2019 Nov 11;8:e52542. doi: 10.7554/eLife.52542. Elife. 2019. PMID: 31710291 Free PMC article.
References
-
- Alilat, M., A. Sivolob, B. Révet, and A. Prunell. 1999. Nucleosome dynamics. Protein and DNA contributions in the chiral transition of the tetrasome, the histone (H3-H4)2 tetramer-DNA particle. J. Mol. Biol. 291:815-841. - PubMed
-
- Bailey, K. A., S. L. Pereira, J. Widom, and J. N. Reeve. 2000. Archaeal histone selection of nucleosome positioning sequences and the procaryotic origin of histone-dependent genome evolution. J. Mol. Biol. 303:25-34. - PubMed
-
- Bailey, K. A., F. Marc, K. Sandman, and J. N. Reeve. 2002. Both DNA and histone fold sequences contribute to archaeal nucleosome stability. J. Biol. Chem. 277:9293-9301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
