

Purinergic and vanilloid receptor activation releases glutamate from separate cranial afferent terminals in nucleus tractus solitarius
- PMID: 15152030
- PMCID: PMC6729471
- DOI: 10.1523/JNEUROSCI.0753-04.2004
Purinergic and vanilloid receptor activation releases glutamate from separate cranial afferent terminals in nucleus tractus solitarius
Abstract
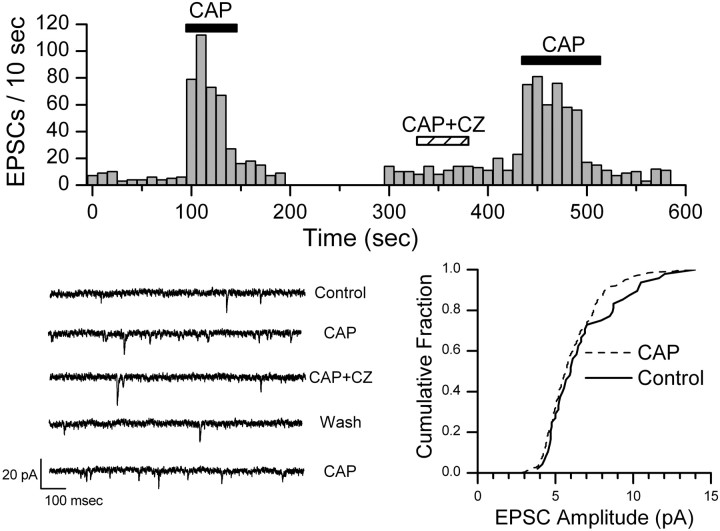
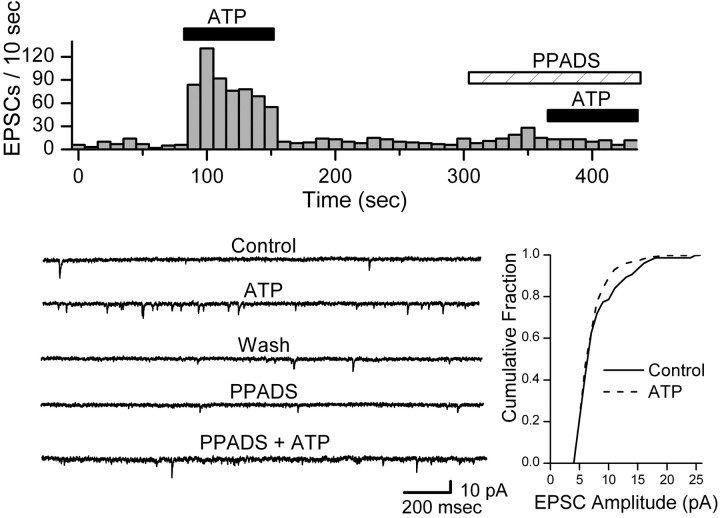
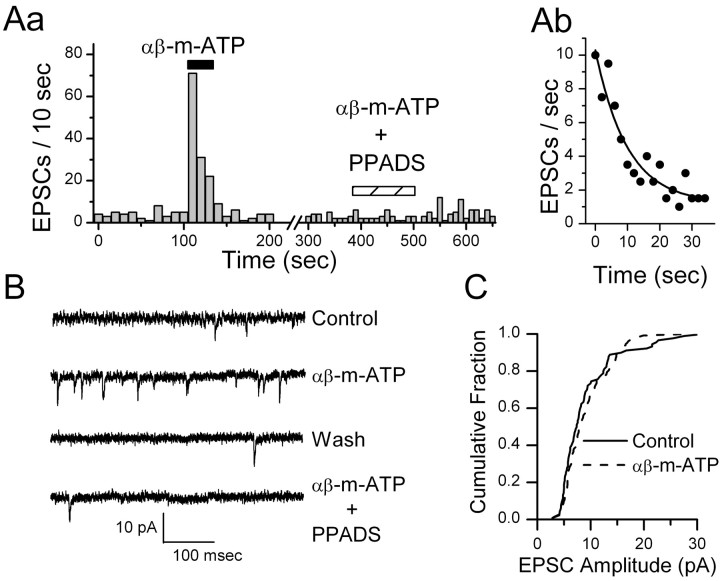
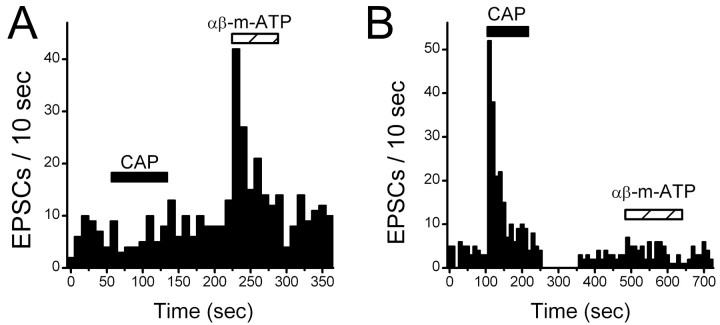
Vanilloid (VR1) and purinergic (P2X) receptors are found in cranial afferent neurons in nodose ganglia and their central terminations within the solitary tract nucleus (NTS), but little is known about their function. We mechanically dissociated dorsomedial NTS neurons to preserve attached native synapses and tested for VR1 and P2X function primarily in spindle-shaped neurons resembling intact second-order neurons. All neurons (n = 95) exhibited spontaneous glutamate (EPSCs) and GABA (IPSCs)-mediated synaptic currents. VR1 agonist capsaicin (CAP; 100 nm) reversibly increased EPSC frequency, effects blocked by capsazepine. ATP (100 microm) increased EPSC frequency, actions blocked by P2X antagonist pyridoxalphosphate-6-azophenyl-2', 4'-disulfonic acid (PPADS; 20 microm). In all CAP-resistant neurons, P2X agonist alphabeta-methylene-ATP (alphabeta-m-ATP) increased EPSC frequency. Neither CAP nor alphabeta-m-ATP altered EPSC amplitudes, kinetics, or holding currents. Thus, activation of VR1 and P2X receptors selectively facilitated presynaptic glutamate release on different NTS neurons. PPADS and 2',3'-O-(2,4,6-trinitrophenyl)-ATP blocked alphabeta-m-ATP responses, but P2X1-selective antagonist NF023 (8,8'-[carbonylbis (imino-3,1-phenylene carbonylimino)]bis-1,3,5-naphthalenetrisulfonic acid) did not. The pharmacological profile and transient kinetics of ATP responses are consistent with P2X3 homomeric receptors. TTX and Cd(2+) did not eliminate agonist-evoked EPSC frequency increases, suggesting that voltage-gated sodium and calcium channels are not required. In nodose ganglia, CAP but not alphabeta-m-ATP evoked inward currents in slow conducting neurons and the converse pattern in myelinated, rapidly conducting neurons (n = 14). Together, results are consistent with segregation of glutamatergic terminals into either P2X sensitive or VR1 sensitive that correspondingly identify myelinated and unmyelinated afferent pathways at the NTS.
Figures



Similar articles
-
Vanilloid receptors presynaptically modulate cranial visceral afferent synaptic transmission in nucleus tractus solitarius.J Neurosci. 2002 Sep 15;22(18):8222-9. doi: 10.1523/JNEUROSCI.22-18-08222.2002. J Neurosci. 2002. PMID: 12223576 Free PMC article.
-
Developmental changes in P2X purinoceptors on glycinergic presynaptic nerve terminals projecting to rat substantia gelatinosa neurones.J Physiol. 2001 Oct 15;536(Pt 2):505-19. doi: 10.1111/j.1469-7793.2001.0505c.xd. J Physiol. 2001. PMID: 11600685 Free PMC article.
-
ATP facilitates glutamatergic neurotransmission to cardiac vagal neurons in the nucleus ambiguus.Brain Res. 2008 Mar 27;1201:88-92. doi: 10.1016/j.brainres.2008.01.065. Epub 2008 Feb 6. Brain Res. 2008. PMID: 18295749 Free PMC article.
-
Contributions of P2X3 homomeric and heteromeric channels to acute and chronic pain.Expert Opin Ther Targets. 2003 Aug;7(4):513-22. doi: 10.1517/14728222.7.4.513. Expert Opin Ther Targets. 2003. PMID: 12885270 Review.
-
P2X receptors in peripheral neurons.Prog Neurobiol. 2001 Oct;65(2):107-34. doi: 10.1016/s0301-0082(01)00005-3. Prog Neurobiol. 2001. PMID: 11403876 Review.
Cited by
-
Of apples and oranges: GABA and glutamate transmission in neurones of the nucleus tractus solitarii could not be more different.J Physiol. 2012 Nov 15;590(22):5559. doi: 10.1113/jphysiol.2012.245340. J Physiol. 2012. PMID: 23154854 Free PMC article. No abstract available.
-
Understanding diverse TRPV1 signaling - an update.F1000Res. 2019 Nov 25;8:F1000 Faculty Rev-1978. doi: 10.12688/f1000research.20795.1. eCollection 2019. F1000Res. 2019. PMID: 31824648 Free PMC article. Review.
-
NMDA receptors control vagal afferent excitability in the nucleus of the solitary tract.Brain Res. 2015 Jan 21;1595:84-91. doi: 10.1016/j.brainres.2014.11.010. Epub 2014 Nov 15. Brain Res. 2015. PMID: 25446446 Free PMC article.
-
Characterization of synapses in the rat subnucleus centralis of the nucleus tractus solitarius.J Neurophysiol. 2015 Jan 15;113(2):466-74. doi: 10.1152/jn.00598.2014. Epub 2014 Oct 29. J Neurophysiol. 2015. PMID: 25355962 Free PMC article.
-
Vasopressin inhibits glutamate release via two distinct modes in the brainstem.J Neurosci. 2006 Jun 7;26(23):6131-42. doi: 10.1523/JNEUROSCI.5176-05.2006. J Neurosci. 2006. PMID: 16763021 Free PMC article.
References
-
- Akaike N, Harata N (1994) Nystatin perforated patch recording and its applications to analyses of intracellular mechanisms. Jpn J Physiol 44: 433–473. - PubMed
-
- Andresen MC, Kunze DL (1994) Nucleus tractus solitarius: gateway to neural circulatory control. Annu Rev Physiol 56: 93–116. - PubMed
-
- Burnstock G (2000) P2X receptors in sensory neurones. Br J Anaesth 84: 476–488. - PubMed
-
- Caterina MJ, Julius D (2001) The vanilloid receptor: a molecular gateway to the pain pathway. Annu Rev Neurosci 24: 487–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous