

Cholecystokinin modulates migration of gonadotropin-releasing hormone-1 neurons
- PMID: 15152034
- PMCID: PMC6729465
- DOI: 10.1523/JNEUROSCI.0649-04.2004
Cholecystokinin modulates migration of gonadotropin-releasing hormone-1 neurons
Abstract
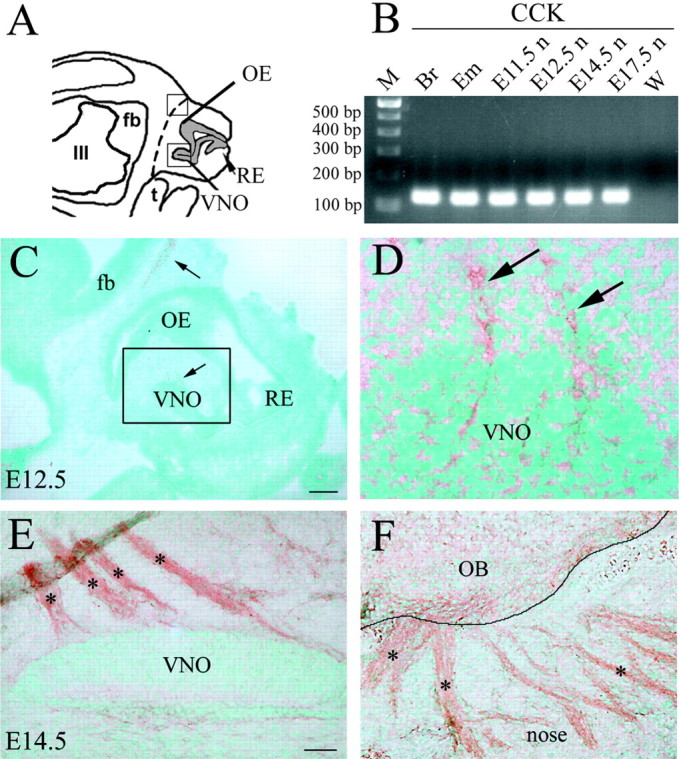
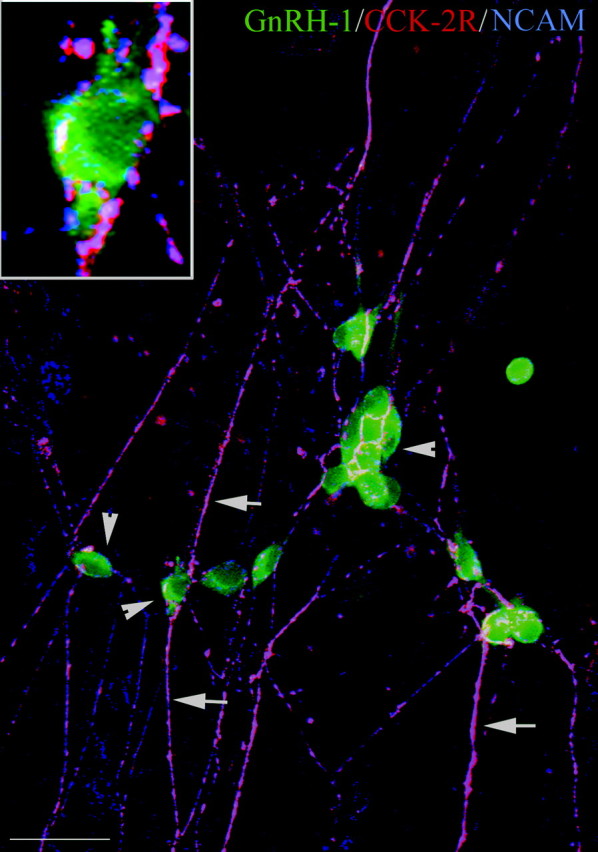
Expression of the brain-gut peptide cholecystokinin (CCK) in the developing olfactory-gonadotropin-releasing hormone-1 (GnRH-1) neuroendocrine systems was characterized, and the function of CCK in these systems was analyzed both in vivo and in vitro. We present novel data demonstrating that CCK transcript and protein are expressed in sensory cells in the developing olfactory epithelium and vomeronasal organ, with both ligand and receptors (CCK-1R and CCK-2R) found on olfactory axons throughout prenatal development. In addition, migrating GnRH-1 neurons in nasal regions express CCK-1R but not CCK-2R receptors. The role of CCK in olfactory-GnRH-1 system development was evaluated using nasal explants, after assessing that the in vivo expression of both CCK and CCK receptors was mimicked in this in vitro model. Exogenous application of CCK (10(-7) m) reduced both olfactory axon outgrowth and migration of GnRH-1 cells. This inhibition was mediated by CCK-1R receptors. Moreover, CCK-1R but not CCK-2R antagonism caused a shift in the location of GnRH-1 neurons, increasing the distance that the cells migrated. GnRH-1 neuronal migration in mice carrying a genetic deletion of either CCK-1R or CCK-2R receptor genes was also analyzed. At embryonic day 14.5, the total number of GnRH-1 cells was identical in wild-type and mutant mice; however, the number of GnRH-1 neurons within forebrain was significantly greater in CCK-1R-/- embryos, consistent with an accelerated migratory process. These results indicate that CCK provides an inhibitory influence on GnRH-1 neuronal migration, contributing to the appropriate entrance of these neuroendocrine cells into the brain, and thus represent the first report of a developmental role for CCK.
Figures






Similar articles
-
Lactosamine modulates the rate of migration of GnRH neurons during mouse development.Eur J Neurosci. 2006 Aug;24(3):654-60. doi: 10.1111/j.1460-9568.2006.04955.x. Eur J Neurosci. 2006. PMID: 16930397
-
Cholecystokinin directly inhibits neuronal activity of primary gonadotropin-releasing hormone cells through cholecystokinin-1 receptor.Endocrinology. 2007 Jan;148(1):63-71. doi: 10.1210/en.2006-0758. Epub 2006 Oct 5. Endocrinology. 2007. PMID: 17023535
-
CXCR4/SDF-1 system modulates development of GnRH-1 neurons and the olfactory system.Dev Neurobiol. 2008 Mar;68(4):487-503. doi: 10.1002/dneu.20594. Dev Neurobiol. 2008. PMID: 18188864
-
Gonadotropin-releasing hormone neuronal migration.Semin Reprod Med. 2007 Sep;25(5):305-12. doi: 10.1055/s-2007-984736. Semin Reprod Med. 2007. PMID: 17710726 Review.
-
Developmental aspect of the gonadotropin-releasing hormone system.Mol Cell Endocrinol. 2001 Dec 20;185(1-2):173-84. doi: 10.1016/s0303-7207(01)00616-5. Mol Cell Endocrinol. 2001. PMID: 11738807 Review.
Cited by
-
Altered ATP7A expression and other compensatory responses in a murine model of Menkes disease.Neurobiol Dis. 2007 Sep;27(3):278-91. doi: 10.1016/j.nbd.2007.05.004. Epub 2007 May 23. Neurobiol Dis. 2007. PMID: 17588765 Free PMC article.
-
Prenatal expression of cholecystokinin (CCK) in the central nervous system (CNS) of mouse.Neurosci Lett. 2008 Jun 13;438(1):96-101. doi: 10.1016/j.neulet.2008.04.042. Epub 2008 May 5. Neurosci Lett. 2008. PMID: 18462884 Free PMC article.
-
A role for FE65 in controlling GnRH-1 neurogenesis.J Neurosci. 2011 Jan 12;31(2):480-91. doi: 10.1523/JNEUROSCI.4698-10.2011. J Neurosci. 2011. PMID: 21228158 Free PMC article.
-
Progesterone directly and rapidly inhibits GnRH neuronal activity via progesterone receptor membrane component 1.Endocrinology. 2012 Sep;153(9):4457-69. doi: 10.1210/en.2012-1122. Epub 2012 Jul 20. Endocrinology. 2012. PMID: 22822163 Free PMC article.
-
Chloride Accumulators NKCC1 and AE2 in Mouse GnRH Neurons: Implications for GABAA Mediated Excitation.PLoS One. 2015 Jun 25;10(6):e0131076. doi: 10.1371/journal.pone.0131076. eCollection 2015. PLoS One. 2015. PMID: 26110920 Free PMC article.
References
-
- Atkinson ME, Shehab SA (1986) Peripheral axotomy of the rat mandibular trigeminal nerve leads to an increase in VIP and decrease of other primary afferent neuropeptides in the spinal trigeminal nucleus. Regul Pept 16: 69–81. - PubMed
-
- Beinfeld MC (1983) Cholecystokinin in the central nervous system: a minireview. Neuropeptides 3: 411–427. - PubMed
-
- Beinfeld MC (2001) An introduction to neuronal cholecystokinin. Peptides 22: 1197–1200. - PubMed
-
- Bless EP, Westaway WA, Schwarting GA, Tobet SA (2000) Effects of gamma-aminobutyric acid(A) receptor manipulation on migrating gonadotropin-releasing hormone neurons through the entire migratory route in vivo and in vitro. Endocrinology 141: 1254–1262. - PubMed
-
- Bock MG, DiPardo RM, Evans BE, Rittle KE, Whitter WL, Veber DF, Anderson PF, Freidinger RM (1989) Benzodiazepin gastrin and brain cholecystokinin receptor ligands: L-365, 260. J Med Chem 32: 13–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases