

A seed for Alzheimer amyloid in the brain
- PMID: 15152051
- PMCID: PMC6729458
- DOI: 10.1523/JNEUROSCI.0861-04.2004
A seed for Alzheimer amyloid in the brain
Abstract
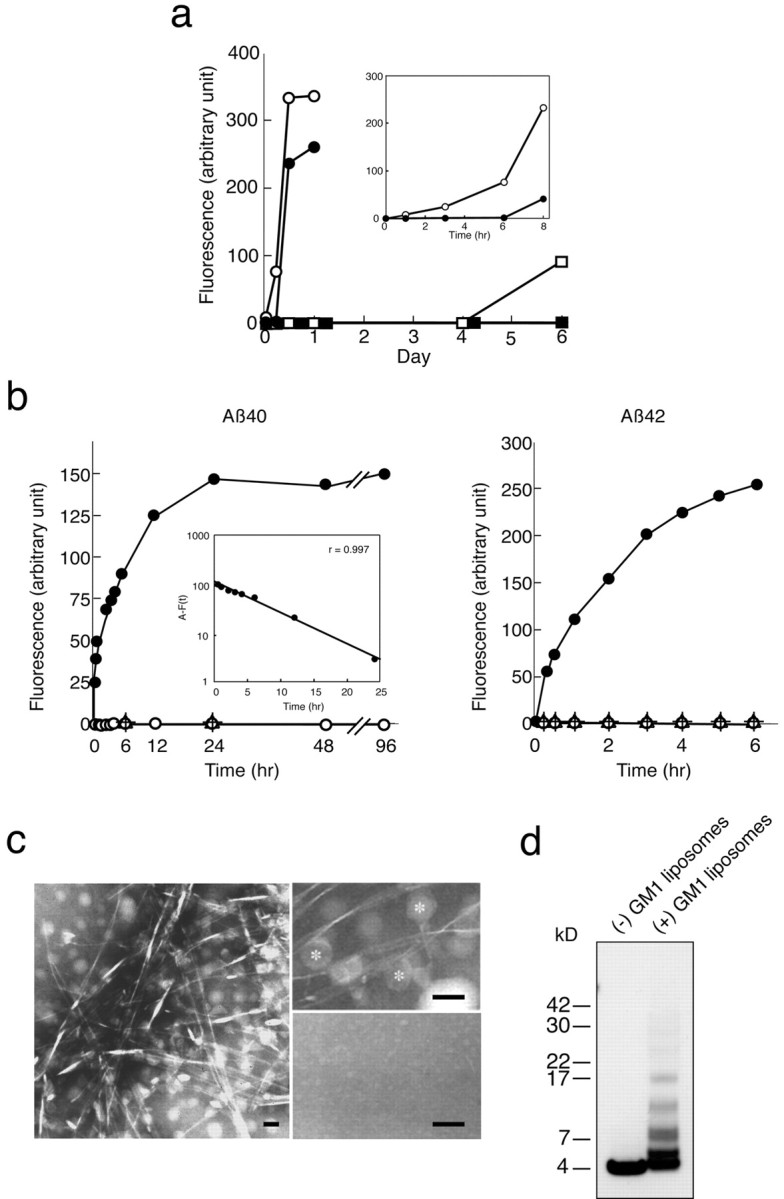
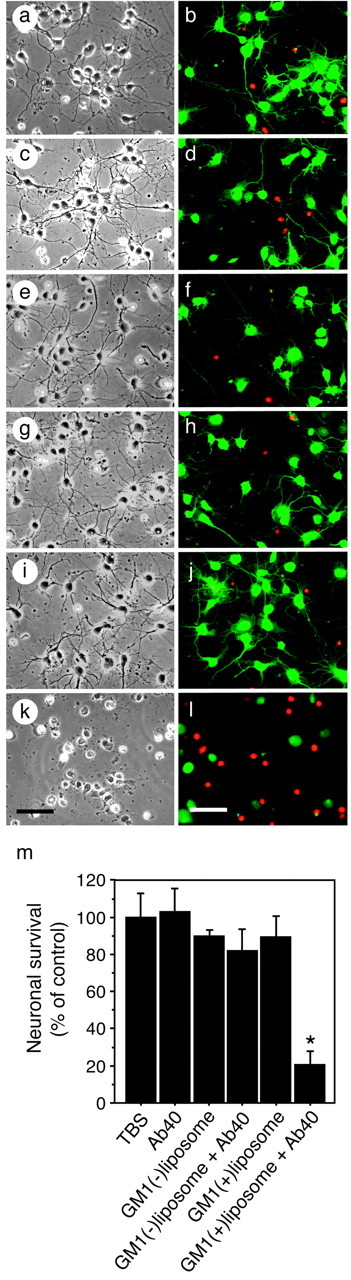
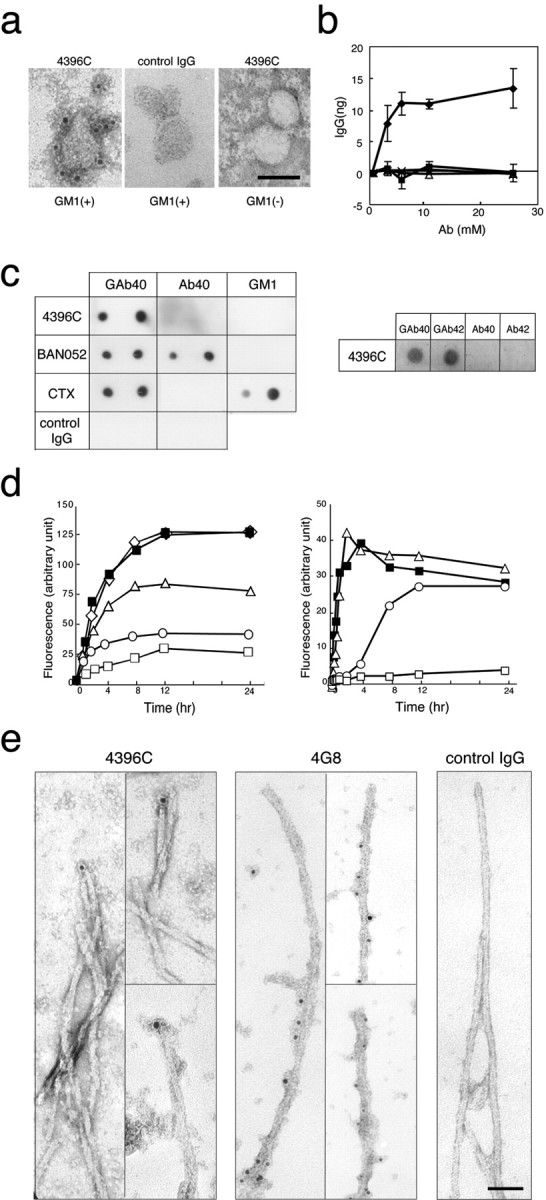
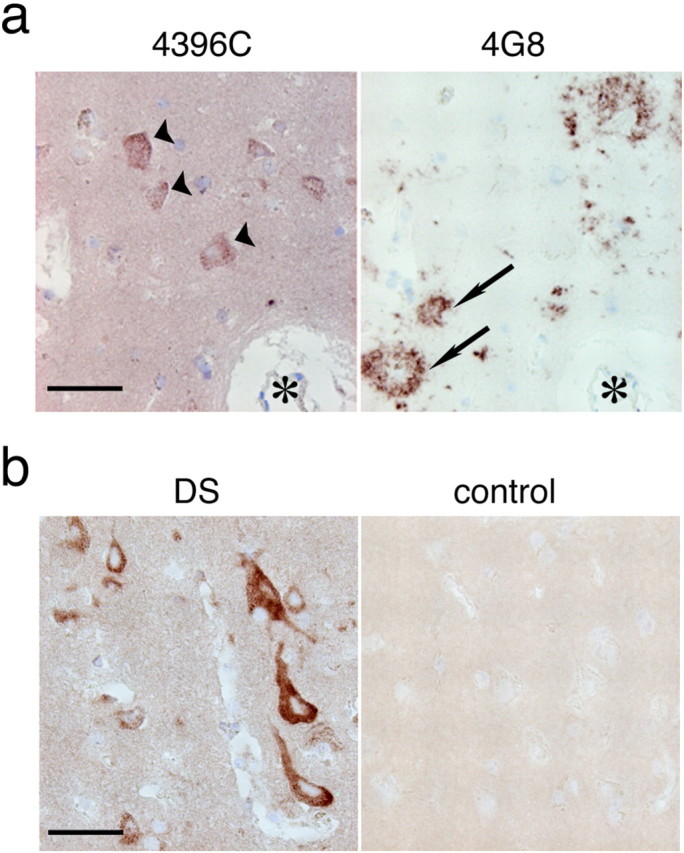
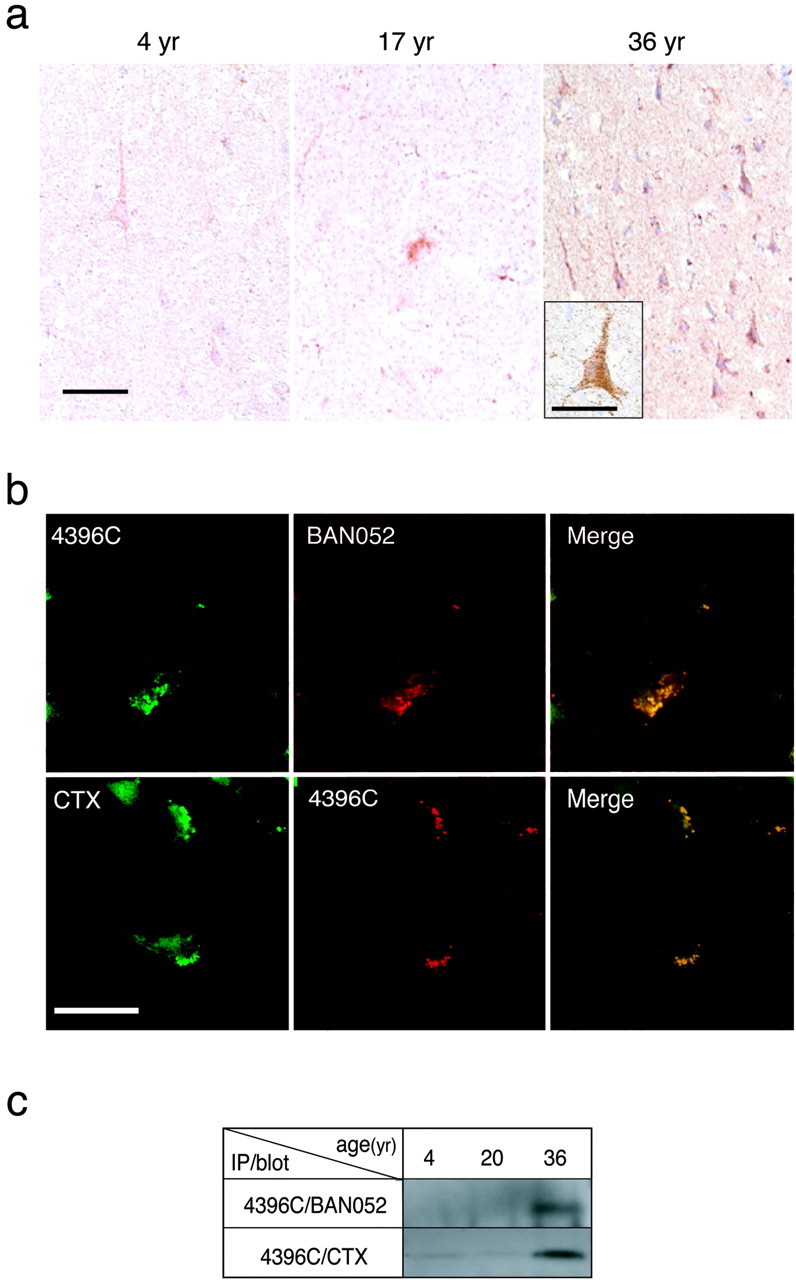
A fundamental question about the early pathogenesis of Alzheimer's disease (AD) concerns how toxic aggregates of amyloid beta protein (Abeta) are formed from its nontoxic soluble form. We hypothesized previously that GM1 ganglioside-bound Abeta (GAbeta) is involved in the process. We now examined this possibility using a novel monoclonal antibody raised against GAbeta purified from an AD brain. Here, we report that GAbeta has a conformation distinct from that of soluble Abeta and initiates Abeta aggregation by acting as a seed. Furthermore, GAbeta generation in the brain was validated by both immunohistochemical and immunoprecipitation studies. These results imply a mechanism underlying the onset of AD and suggest that an endogenous seed can be a target of therapeutic strategy.
Figures
Similar articles
-
GM1 ganglioside and the seeding of amyloid in Alzheimer's disease: endogenous seed for Alzheimer amyloid.Neuroscientist. 2005 Jun;11(3):250-60. doi: 10.1177/1073858405275177. Neuroscientist. 2005. PMID: 15911874 Review.
-
Role of gangliosides in Alzheimer's disease.Biochim Biophys Acta. 2007 Aug;1768(8):1943-51. doi: 10.1016/j.bbamem.2007.01.018. Epub 2007 Jan 28. Biochim Biophys Acta. 2007. PMID: 17321494 Review.
-
How do membranes initiate Alzheimer's Disease? Formation of toxic amyloid fibrils by the amyloid β-protein on ganglioside clusters.Acc Chem Res. 2014 Aug 19;47(8):2397-404. doi: 10.1021/ar500127z. Epub 2014 Jul 16. Acc Chem Res. 2014. PMID: 25029558
-
Interactions of amyloid beta-protein with various gangliosides in raft-like membranes: importance of GM1 ganglioside-bound form as an endogenous seed for Alzheimer amyloid.Biochemistry. 2002 Jun 11;41(23):7385-90. doi: 10.1021/bi0255874. Biochemistry. 2002. PMID: 12044171
-
Abeta polymerization through interaction with membrane gangliosides.Biochim Biophys Acta. 2010 Aug;1801(8):868-77. doi: 10.1016/j.bbalip.2010.01.008. Epub 2010 Feb 1. Biochim Biophys Acta. 2010. PMID: 20117237 Review.
Cited by
-
Change of dynamics of raft-model membrane induced by amyloid-β protein binding.Eur Phys J E Soft Matter. 2013 Jul;36(7):74. doi: 10.1140/epje/i2013-13074-3. Epub 2013 Jul 16. Eur Phys J E Soft Matter. 2013. PMID: 23852578
-
Furin-mediated cleavage of LRP1 and increase in ICD of LRP1 after cerebral ischemia and after exposure of cultured neurons to NMDA.Sci Rep. 2019 Aug 13;9(1):11782. doi: 10.1038/s41598-019-48279-x. Sci Rep. 2019. PMID: 31409872 Free PMC article.
-
Deregulated sphingolipid metabolism and membrane organization in neurodegenerative disorders.Mol Neurobiol. 2010 Jun;41(2-3):314-40. doi: 10.1007/s12035-009-8096-6. Epub 2010 Feb 3. Mol Neurobiol. 2010. PMID: 20127207 Review.
-
The Double-Layered Structure of Amyloid-β Assemblage on GM1-Containing Membranes Catalytically Promotes Fibrillization.ACS Chem Neurosci. 2023 Aug 2;14(15):2648-2657. doi: 10.1021/acschemneuro.3c00192. Epub 2023 Jul 23. ACS Chem Neurosci. 2023. PMID: 37482658 Free PMC article.
-
Evidence that small molecule enhancement of β-hexosaminidase activity corrects the behavioral phenotype in Dutch APP(E693Q) mice through reduction of ganglioside-bound Aβ.Mol Psychiatry. 2015 Feb;20(1):109-17. doi: 10.1038/mp.2014.135. Epub 2014 Oct 28. Mol Psychiatry. 2015. PMID: 25349165 Free PMC article.
References
-
- Ariga T, Kobayashi K, Hasegawa A, Kiso M, Ishida H, Miyatake T (2001) Characterization of high-affinity binding between gangliosides and amyloid beta-protein. Arch Biochem Biophys 388: 225–230. - PubMed
-
- Bard F, Cannon C, Barbour R, Burke RL, Games D, Grajeda H, Guido T, Hu K, Huang J, Johnson-Wood K, Khan K, Kholodenko D, Lee M, Lieberburg I, Motter R, Nguyen M, Soriano F, Vasquez N, Weiss K, Welch B, et al. (2000) Peripherally administered antibodies against amyloid beta-peptide enter the central nervous system and reduce pathology in a mouse model of Alzheimer disease. Nat Med 6: 916–919. - PubMed
-
- Barrett JE, Wells DC, Conrad GW (1999) Pretreatment methods to improve nerve immunostaining in corneas from long-term fixed embryonic quail eyes. J Neurosci Methods 92: 161–168. - PubMed
-
- Binding MM, Jones ST (1996) Rodent to human antibodies by CDR grafting. In: Antibody engineering (Mcmafferty J, Hoogenboon HR, Chiswell DJ, eds), pp 147–168. Oxford: Oxford UP.
-
- Boon ME, Kok LP (1991) Formalin is deleterious to cytoskeleton proteins: do we need to replace it by formalin-free Kryofix? Eur J Morphol 29: 173–180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical