

Mechanism of action at the molecular level of the antiviral drug 3(2H)-isoflavene against type 2 poliovirus
- PMID: 15155227
- PMCID: PMC415587
- DOI: 10.1128/AAC.48.6.2233-2243.2004
Mechanism of action at the molecular level of the antiviral drug 3(2H)-isoflavene against type 2 poliovirus
Abstract
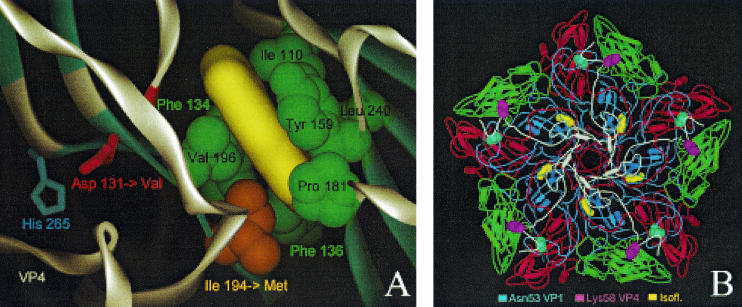
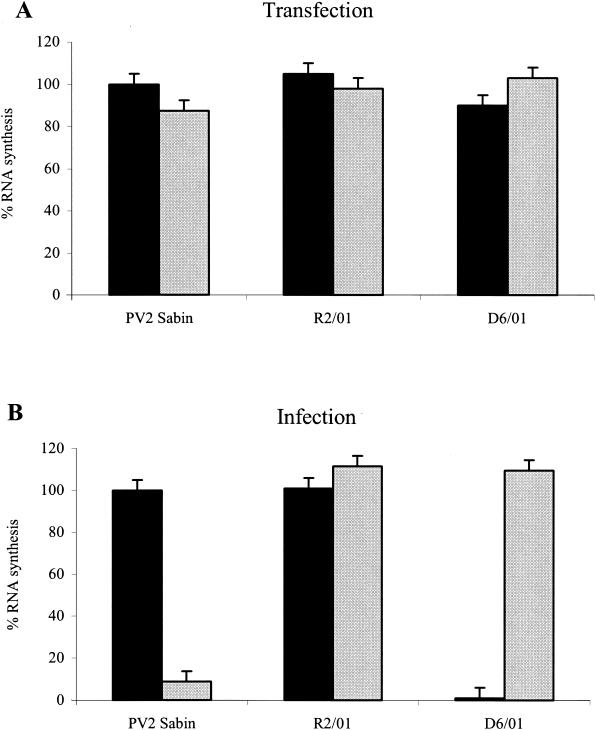
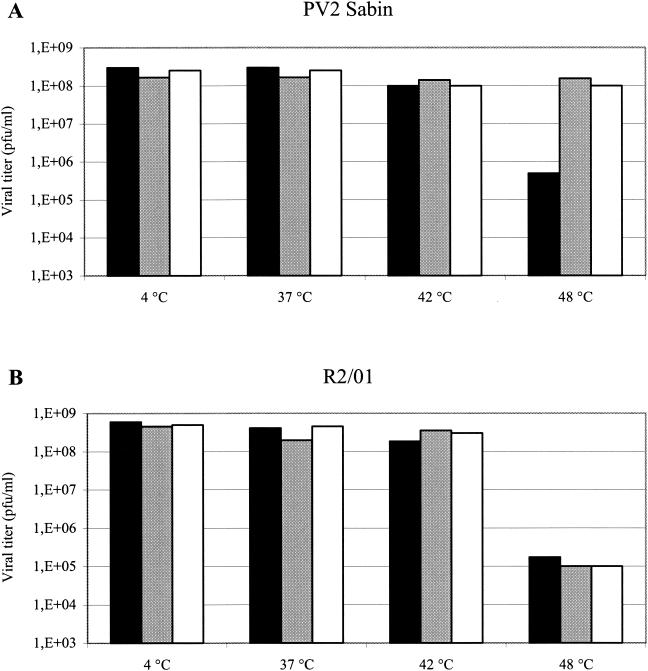
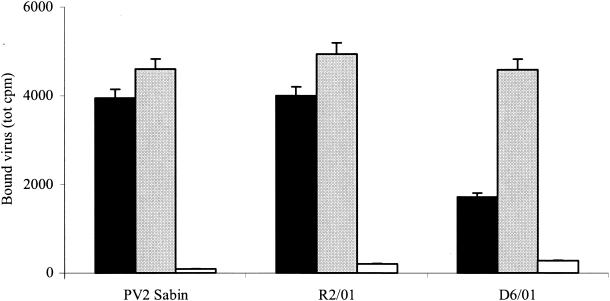
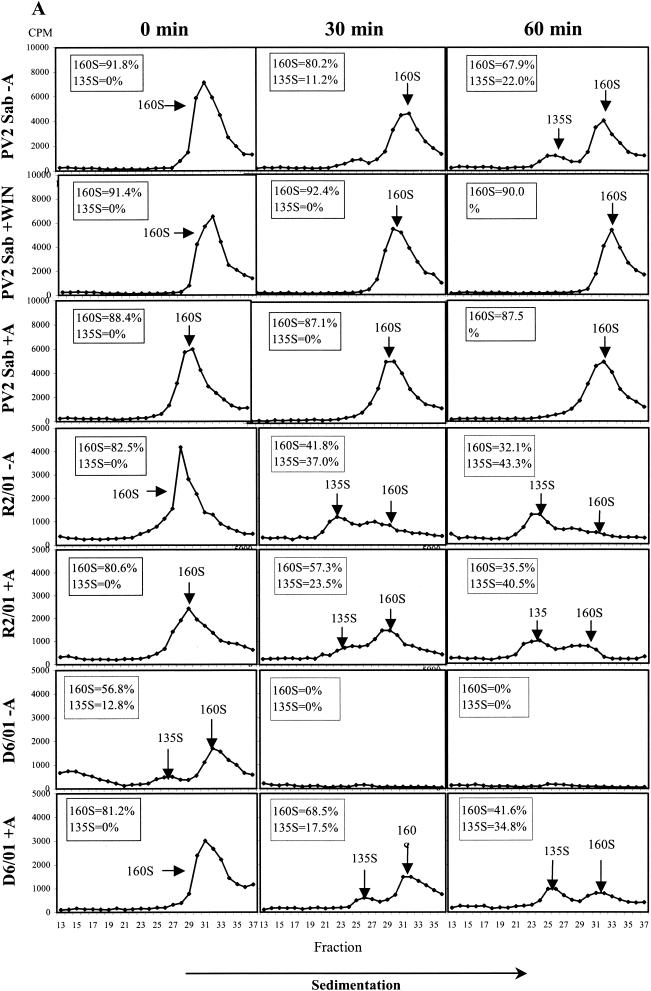
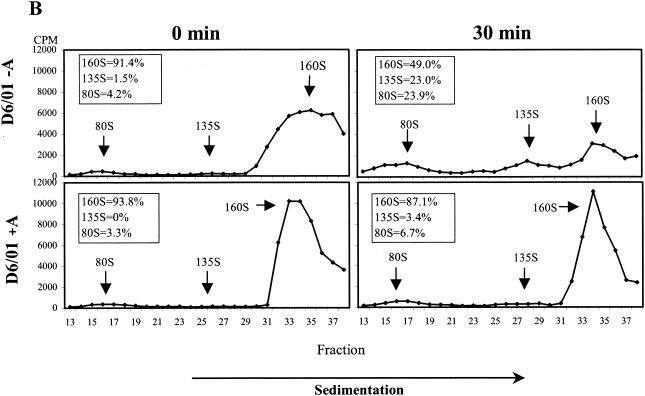
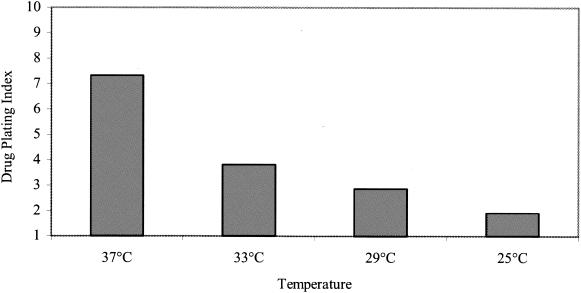
The mechanism of action of the antiviral compound 3(2H)-isoflavene against Sabin type 2 poliovirus has been studied, and interference with virus uncoating was demonstrated. Isolation and sequencing of drug-resistant variants revealed single amino acid substitutions (I194M or D131V) in the VP1 capsid protein. While M194 is located in a hydrophobic pocket and should partially fill the space occupied by the isoflavene ring, V131 is exposed on the VP1 surface, forming a contact with VP4. The D131V mutation most likely induces local conformational changes in VP1 and/or VP4 that affect viral flexibility. Two dependent variants, N53S of VP1 and K58E of VP4, both located on the inner surface of the capsid, near the threefold axis of symmetry, were also selected. Both mutations affected viral stability, allowing the transition to 135S particles in the absence of drug, without the involvement of the viral receptor.
Figures
Similar articles
-
Isolation and biological characterization of 3(2H)-isoflavene-resistant and -dependent poliovirus type 2 Sabin mutants.J Gen Virol. 1999 Jan;80 ( Pt 1):157-167. doi: 10.1099/0022-1317-80-1-157. J Gen Virol. 1999. PMID: 9934698
-
H1PVAT is a novel and potent early-stage inhibitor of poliovirus replication that targets VP1.Antiviral Res. 2014 Oct;110:1-9. doi: 10.1016/j.antiviral.2014.07.003. Epub 2014 Jul 17. Antiviral Res. 2014. PMID: 25043639
-
Distribution of drug resistance mutations in type 3 poliovirus identifies three regions involved in uncoating functions.J Virol. 1994 Dec;68(12):8193-201. doi: 10.1128/JVI.68.12.8193-8201.1994. J Virol. 1994. PMID: 7966611 Free PMC article.
-
Early events in poliovirus infection: virus-receptor interactions.Proc Natl Acad Sci U S A. 1996 Oct 15;93(21):11378-81. doi: 10.1073/pnas.93.21.11378. Proc Natl Acad Sci U S A. 1996. PMID: 8876143 Free PMC article. Review.
-
My Cousin, My Enemy: quasispecies suppression of drug resistance.Curr Opin Virol. 2016 Oct;20:106-111. doi: 10.1016/j.coviro.2016.09.011. Epub 2016 Oct 17. Curr Opin Virol. 2016. PMID: 27764731 Free PMC article. Review.
Cited by
-
Viral adaptation to an antiviral protein enhances the fitness level to above that of the uninhibited wild type.J Virol. 2009 Nov;83(22):11746-50. doi: 10.1128/JVI.01297-09. Epub 2009 Sep 2. J Virol. 2009. PMID: 19726521 Free PMC article.
-
Modelling and targeting mitochondrial protein tyrosine phosphatase 1: a computational approach.In Silico Pharmacol. 2022 Jan 17;10(1):3. doi: 10.1007/s40203-022-00119-z. eCollection 2022. In Silico Pharmacol. 2022. PMID: 35111562 Free PMC article.
-
Mechanism of action and capsid-stabilizing properties of VHHs with an in vitro antipolioviral activity.J Virol. 2014 Apr;88(8):4403-13. doi: 10.1128/JVI.03402-13. Epub 2014 Feb 5. J Virol. 2014. PMID: 24501405 Free PMC article.
-
Antiviral Ability of Kalanchoe gracilis Leaf Extract against Enterovirus 71 and Coxsackievirus A16.Evid Based Complement Alternat Med. 2012;2012:503165. doi: 10.1155/2012/503165. Epub 2012 May 15. Evid Based Complement Alternat Med. 2012. PMID: 22666293 Free PMC article.
-
Potent antiviral agents fail to elicit genetically-stable resistance mutations in either enterovirus 71 or Coxsackievirus A16.Antiviral Res. 2015 Dec;124:77-82. doi: 10.1016/j.antiviral.2015.10.006. Epub 2015 Oct 30. Antiviral Res. 2015. PMID: 26522770 Free PMC article.
References
-
- Bayer, N., E. Prchla, M. Schwab, D. Blaas, and R. Fuchs. 1999. Human rhinovirus HRV14 uncoats from early endosomes in the presence of bafilomycin. FEBS Lett. 463:175-178. - PubMed
-
- Burali, C., N. Desideri, M. L. Stein, C. Conti, and N. Orsi. 1987. Synthesis and anti-rhinovirus activity of halogen-substituted isoflavenes and isoflavanes. Eur. J. Med. Chem. 22:119-123.
-
- Caliguiri, L. A., J. J. McSharry, and G. W. Lawrence. 1980. Effect of arildone on modifications of poliovirus in vitro. Virology 105:86-93. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
