

Review
doi: 10.1128/IAI.72.6.3089-3096.2004.
Model for immune responses to Mycobacterium avium subspecies paratuberculosis in cattle
Affiliations
- PMID: 15155609
- PMCID: PMC415675
- DOI: 10.1128/IAI.72.6.3089-3096.2004
Item in Clipboard
Review
Model for immune responses to Mycobacterium avium subspecies paratuberculosis in cattle
Infect Immun.
2004 Jun.
No abstract available
Figures
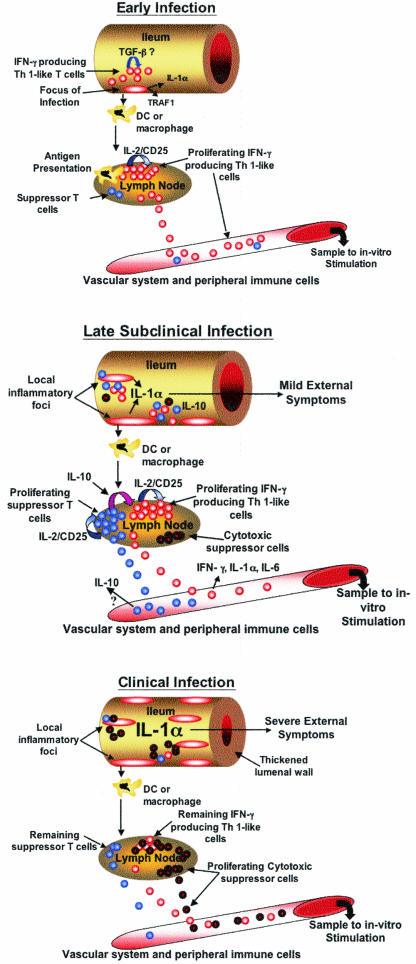
Proposed in vivo immune responses during progressive infection with M. avium subsp. paratuberculosis. (A) Early lesions associated with M. avium subsp. paratuberculosis infection likely develop in ileal Peyer's patches and consist of numerous infected macrophages. Infected macrophages express enhanced amounts of adhesion molecules, IL-1α,and TRAF1, leading to enhanced macrophage survival. Dendritic cells (DC) or macrophages successful in degrading M. avium subsp. paratuberculosis migrate to local lymph nodes, presenting antigen and stimulating reactive T cells to produce IFN-γ and proliferate via IL-2/CD25 signaling (red circles). IFN-γ-producing proinflamma-tory T cells released to the vascular system circulate, eventually migrating to sites of M. avium subsp. paratuberculosis infection, perhaps attracted by enhanced levels of IL-8 produced by infected macrophages. Both antigen-primed CD4+ T cells and infected macrophages produce IFN-γ, possibly activating newly recruited macrophages and helping to control or restrict the infection. A suppressor cell population capable of producing IL-10 is likely already present in lymph nodes, albeit in very small numbers (blue circles). At sites of infection, transforming growth factor β (TGF-β) may be expressed in a attempt to suppress or dampen the early proinflammatory response. (B) In late subclinical infection, continued expansion of the infection has occurred through migration of infected macrophages throughout the ileum. Failure to express TNF-α and to contain the infection in granulomas is one factor likely contributing to this disseminated infection. In late subclinical infection, the numerous macrophages within lesions produce significant amounts of IL-1α, perhaps causing mild IL-1α toxicity. Continued migration of macrophages and/or dendritic cells to local lymph nodes, combined with a prolonged proinflammatory response, has resulted in enhanced proliferation of an IL-10-producing suppressor cell population (blue circles). The balance of proinflammatory (red circles) and suppressor cells leads to loss of IFN-γ and IL-6 production at sites of infection and in local lymph nodes. Reduced production of IFN-γ may help propagate and expand the infection, as newly recruited macrophages would not be sufficiently activated to kill M. avium subsp. paratuberculosis. A population of cytotoxic suppressor cells has begun to proliferate within local lymph nodes (black-outlined red circles), although not in sufficient numbers to control the activated proinflammatory cells. (C) In clinically infected cattle, the ileum is heavily infected and is typically thickened, reducing transport of nutrients. It is proposed that large numbers of infected macrophages produce sufficient IL-1α to cause outward signs of toxicity. In local lymph nodes, continued expansion of a cytotoxic suppressor cell population (black-outlined red circles) has removed many of the M. avium subsp. paratuberculosis-reactive proinflammatory T cells (red circles) present during late subclinical infection. Loss of proinflammatory cells has reduced the need for IL-10-producing suppressor cells (blue circles). Indeed, if these cells are T regulatory cells, their continued proliferation may depend upon IL-2 released from the proinflammatory cells. Complete or nearly complete loss of an antigen-specific response allows the infection with M. avium subsp. paratuberculosis to progress essentially unchecked. Although not represented in this figure, antigen-reactive B-cell populations have continued to proliferate, and the remaining immune response is largely production of IgG1.
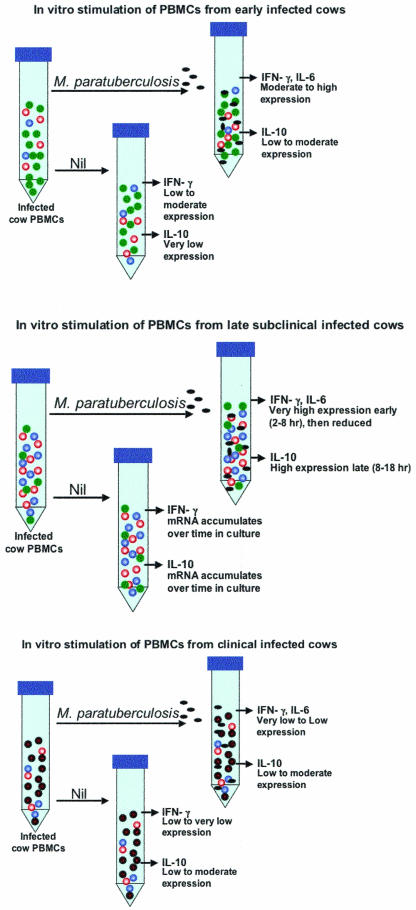
Proposed in vitro responses of PBMCs from progressively infected cows to stimulation with M. avium subsp. paratuberculosis. (A) Removal of peripheral immune cells from early-stage infected cows and stimulation in vitro with M. avium subsp. paratuberculosis results in a minor population of antigen-reactive proinflammatory T cells (red circles) producing significant amounts of IFN-γ. Expression of IL-6 and IL-1α is also upregulated in these proinflammatory cells. A small number of suppressor T cells (blue circles) may also be present in PBMCs from cattle in early stages of infection, but in numbers insufficient to control the proinflammatory response. Many peripheral immune cells are present in PBMC preparations that are not reactive to M. avium subsp. paratuberculosis antigens (green circles). Thus, the predominant response to in vitro stimulation of PBMCs from cattle at this stage of infection is production of IFN-γ, IL-6, and IL-1α.(B) PBMCs during the late subclinical stage of infection contain high numbers of M. avium subsp. paratuberculosis-reactive proinflammatory cells (red circles) that are already activated and produce significant amounts of IFN-γ and IL-6, even without further antigen stimulation in vitro (Nil). In vitro, production of IFN-γ, IL-6, and perhaps IL-1α is rapidly enhanced by introduction of M. avium subsp. paratuberculosis antigens, although this response is eventually quelled by IL-10 produced from large numbers of activated suppressor cells (blue circles). Overall, the percentage of non-M. avium subsp. paratuberculosis-reactive cells (green circles) has been reduced, although the balance of CD4+, CD8+, and γδ TCR+ T cells has not changed appreciably. (C) Due to the presence of large numbers of cytotoxic suppressor cells and reduced numbers of proinflammatory cells, PBMCs extracted during clinical stages of infection produce less IFN-γ and IL-6 in response to M. avium subsp. paratuberculosis antigens than cells from late subclinically infected cattle. Loss of IL-10-producing suppressor cells, which are present in large numbers during subclinical infection, also leads to lower production of this cytokine in PBMCs from clinically infected cattle. Prolonged in vitro stimulation (>16 h) of PBMCs from clinically infected cattle with M. avium subsp. paratuberculosis antigens results in apoptosis of many proinflammatory and cytotoxic suppressor cells. PBMCs from cattle in clinical stages of Johne's disease thus produce little IFN-γ, IL-6, and IL-1α in response to M. avium subsp. paratuberculosis antigens, although continued proliferation of the cytotoxic cell population has caused a profound change in the pattern of gene expression observed in PBMCs from these animals relative to those from uninfected cattle or cattle in earlier stages of infection. Although not shown in this figure, the predominant remaining response to M. avium subsp. paratuberculosis in clinically infected cattle is production of IgG1 from antigen-stimulated B cells. The proposed cytotoxic cell population may also kill non-M. avium subsp. paratuberculosis-reactive T cells in vitro (and perhaps in vivo), thus leading to an observed general immune cell anergy in PBMCs from clinically infected cattle.
Similar articles
-
Secreted antigens of Mycobacterium avium subspecies paratuberculosis as prominent immune targets.Vet Microbiol. 2006 May 31;114(3-4):337-44. doi: 10.1016/j.vetmic.2005.12.005. Epub 2006 Jan 18. Vet Microbiol. 2006. PMID: 16413703
-
Host responses to persistent Mycobacterium avium subspecies paratuberculosis infection in surgically isolated bovine ileal segments.Clin Vaccine Immunol. 2013 Feb;20(2):156-65. doi: 10.1128/CVI.00496-12. Epub 2012 Dec 5. Clin Vaccine Immunol. 2013. PMID: 23221000 Free PMC article.
-
Mycobacterium avium subspecies paratuberculosis infection of cattle does not diminish peripheral blood-derived macrophage mycobactericidal activity.Immunol Lett. 2006 Sep 15;107(1):76-9. doi: 10.1016/j.imlet.2006.06.004. Epub 2006 Jul 7. Immunol Lett. 2006. PMID: 16884783
-
[Pathogenesis and immune reactions of paratuberculosis].Dtsch Tierarztl Wochenschr. 2002 Dec;109(12):507-9. Dtsch Tierarztl Wochenschr. 2002. PMID: 12596563 Review. German.
-
Mycobacterium paratuberculosis and the bovine immune system.Anim Health Res Rev. 2001 Dec;2(2):141-61. Anim Health Res Rev. 2001. PMID: 11831436 Review.
Cited by
-
Bovine Neutrophils Release Extracellular Traps and Cooperate With Macrophages in Mycobacterium avium subsp. paratuberculosis clearance In Vitro.Front Immunol. 2021 Mar 17;12:645304. doi: 10.3389/fimmu.2021.645304. eCollection 2021. Front Immunol. 2021. PMID: 33815401 Free PMC article.
-
Pan-genomic analysis of bovine monocyte-derived macrophage gene expression in response to in vitro infection with Mycobacterium avium subspecies paratuberculosis.Vet Res. 2012 Mar 28;43(1):25. doi: 10.1186/1297-9716-43-25. Vet Res. 2012. PMID: 22455317 Free PMC article.
-
Differential cytokine gene expression profiles in the three pathological forms of sheep paratuberculosis.BMC Vet Res. 2007 Aug 14;3:18. doi: 10.1186/1746-6148-3-18. BMC Vet Res. 2007. PMID: 17697353 Free PMC article.
-
Development and Validation of a Novel ELISA for the Specific Detection of Antibodies against Mycobacterium avium Subspecies paratuberculosis Based on a Chimeric Polyprotein.Vet Med Int. 2021 Dec 29;2021:7336848. doi: 10.1155/2021/7336848. eCollection 2021. Vet Med Int. 2021. PMID: 35003619 Free PMC article.
-
Prostaglandin E2 Induction Suppresses the Th1 Immune Responses in Cattle with Johne's Disease.Infect Immun. 2018 Apr 23;86(5):e00910-17. doi: 10.1128/IAI.00910-17. Print 2018 May. Infect Immun. 2018. PMID: 29483289 Free PMC article.
References
-
- Appelberg, R. 1994. Protective role of interferon gamma, tumor necrosis factor alpha and interleukin-6 in Mycobacterium tuberculosis and M. avium infections. Immunobiology 191:520-525. - PubMed
-
- Bannantine, J. P., J. F. Huntley, E. Miltner, J. R. Stabel, and L. E. Bermudez. 2003. The Mycobacterium avium subsp. paratuberculosis 35 kDa protein plays a role in invasion of bovine epithelial cells. Microbiology 149:2061-2069. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
