

Repression of AGAMOUS by BELLRINGER in floral and inflorescence meristems
- PMID: 15155890
- PMCID: PMC490040
- DOI: 10.1105/tpc.021147
Repression of AGAMOUS by BELLRINGER in floral and inflorescence meristems
Abstract
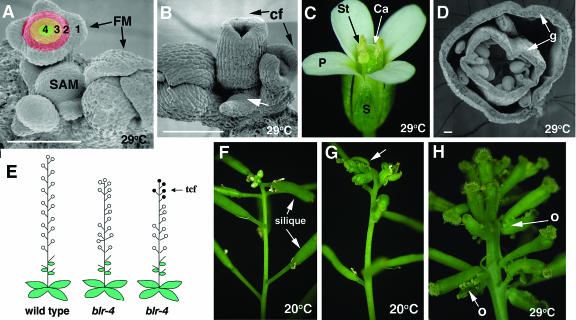
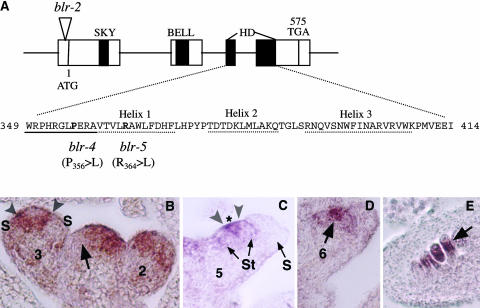
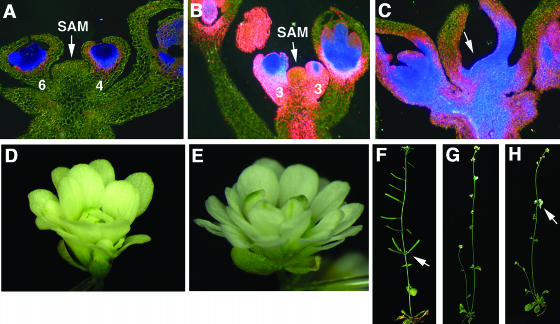
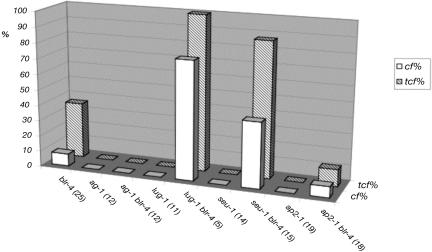
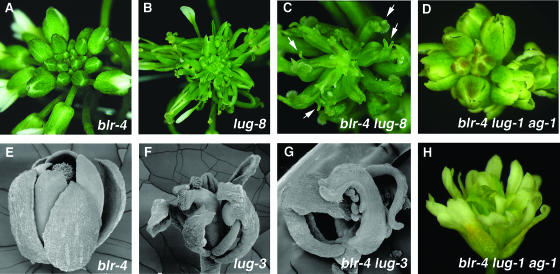
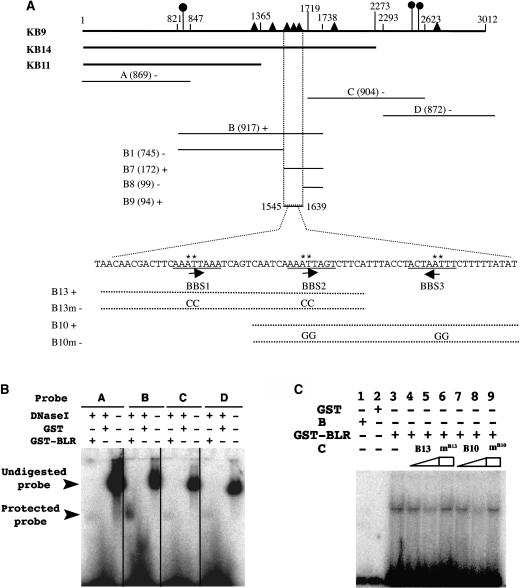
A common aspect of gene regulation in all developmental systems is the sustained repression of key regulatory genes in inappropriate spatial or temporal domains. To understand the mechanism of transcriptional repression of the floral homeotic gene AGAMOUS (AG), we identified two mutations in the BELLRINGER (BLR) gene based on a striking floral phenotype, in which homeotic transformations from sepals to carpels are found in flowers derived from old terminating shoots. Furthermore, this phenotype is drastically enhanced by growth at a high temperature and by combining blr with mutants of LEUNIG and SEUSS, two putative transcriptional corepressors of AG. We showed that the floral phenotype of blr mutants is caused by derepression of AG, suggesting that BLR functions as a transcription repressor. Because BLR encodes a BELL1-like (BELL) homeobox protein, direct binding of BLR to AG cis-regulatory elements was tested by gel-shift assays, and putative BLR binding motifs were identified. In addition, these putative BLR binding motifs were shown to be conserved in 17 of the 29 Brassicaceae species by phylogenetic footprinting. Because BELL homeobox proteins are a family of plant-specific transcription factors with 12 members in Arabidopsis thaliana, our findings will facilitate the identification of regulatory targets of other BELL proteins and help determine their biological functions. The age-dependent and high temperature-enhanced derepression of AG in blr mutants led us to propose that AG expression might be regulated by a thermal time-dependent molecular mechanism.
Copyright 2004 American Society of Plant Biologists
Figures

References
-
- Barolo, S., and Posakony, J.W. (2002). Three habits of highly effective signaling pathways: Principles of transcriptional control by developmental cell signaling. Genes Dev. 16, 1167–1181. - PubMed
-
- Baurle, I., and Laux, T. (2003). Apical meristems: The plant's fountain of youth. Bioessays 25, 961–970. - PubMed
-
- Blazquez, M.A., Soowal, L.N., Lee, I., and Weigel, D. (1997). LEAFY expression and flower initiation in Arabidopsis. Development 124, 3835–3844. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
