

Two Creutzfeldt-Jakob disease agents reproduce prion protein-independent identities in cell cultures
- PMID: 15161970
- PMCID: PMC423270
- DOI: 10.1073/pnas.0400158101
Two Creutzfeldt-Jakob disease agents reproduce prion protein-independent identities in cell cultures
Abstract
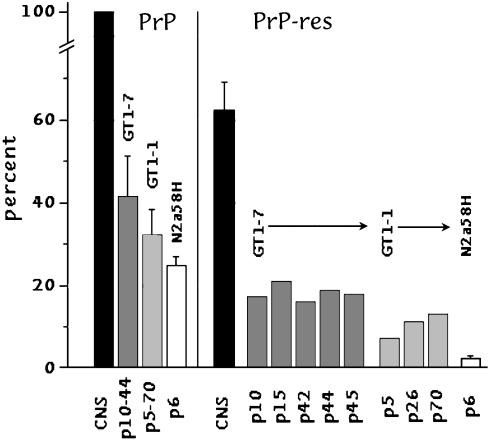
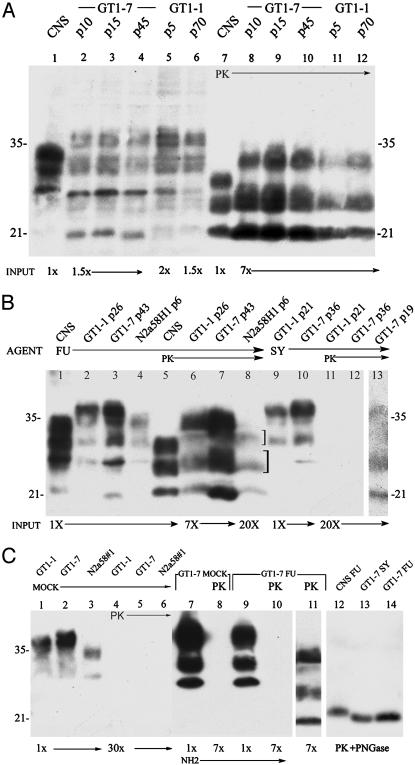
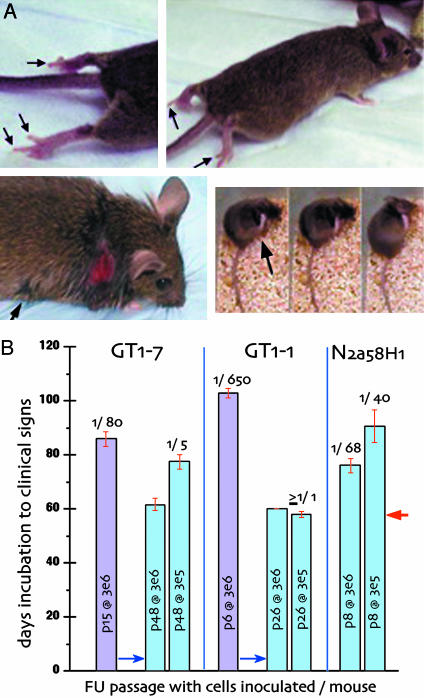
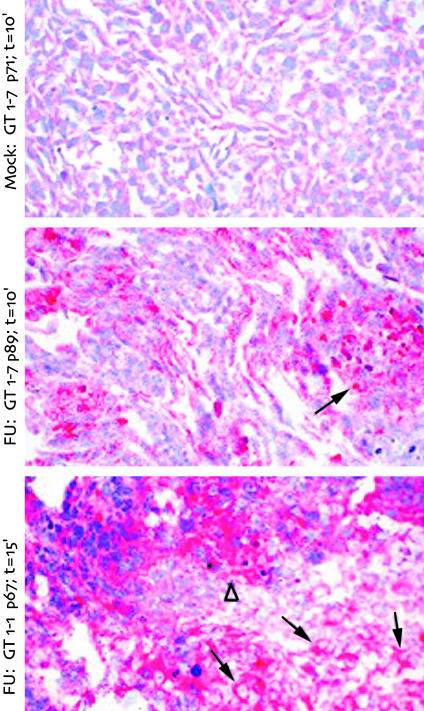
Human Creutzfeldt-Jakob disease (CJD) and similar neurodegenerative diseases such as sheep scrapie are caused by a variety of related infectious agents. They are associated with abnormal host prion protein (PrP), which is assessed by limited proteolysis to yield resistant PrP bands (PrP-res). Although PrP-res has been posited as the infectious agent, purified PrP-res itself is not infectious. To establish the independence of CJD agent characteristics from those of PrP-res, two different mouse-passaged CJD strains were propagated in neuronal cell lines whose PrP-res patterns differ markedly from each other and from those found in infected brain. In mouse brain, the fast CJD strain, FU, elicits many PrP-res deposits, whereas the slow SY strain elicits few. Both strains evoked PrP-res in cultured murine cells, although SY induced PrP-res only transiently. PrP-res patterns in FU- and SY-infected GT1 cells were identical, and were significantly different from those in brain and in N2a cells. Nevertheless, all FU-infected cell lines reproduced their original fast disease in mice, even after extensive subculture, whereas SY-infected cells produced only slow disease. These data indicate PrP-res neither encodes nor alters agent-specific characteristics. PrP-res was also a poor predictor of infectivity because SY cells that had lost PrP-res were approximately 10-fold more infectious than PrP-res-positive cultures. Furthermore, FU titers increased 650-fold, whereas PrP-res remained constant. Passaged FU-infected cells had titers comparable to brain, and >30% of cells displayed abundant cytoplasmic PrP-res aggregates that may trap agent. The continuous substantial replication of CJD in monotypic cells will further the discrimination of agent-specific molecules from pathological host responses to infection.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
