

doi: 10.1113/jphysiol.2004.066001.
Epub 2004 May 28.
Knockout of the ASIC2 channel in mice does not impair cutaneous mechanosensation, visceral mechanonociception and hearing
Affiliations
- PMID: 15169849
- PMCID: PMC1664970
- DOI: 10.1113/jphysiol.2004.066001
Item in Clipboard
Knockout of the ASIC2 channel in mice does not impair cutaneous mechanosensation, visceral mechanonociception and hearing
J Physiol.
.
Abstract
Mechanosensitive cation channels are thought to be crucial for different aspects of mechanoperception, such as hearing and touch sensation. In the nematode C. elegans, the degenerins MEC-4 and MEC-10 are involved in mechanosensation and were proposed to form mechanosensitive cation channels. Mammalian degenerin homologues, the H(+)-gated ASIC channels, are expressed in sensory neurones and are therefore interesting candidates for mammalian mechanosensors. We investigated the effect of an ASIC2 gene knockout in mice on hearing and on cutaneous mechanosensation and visceral mechanonociception. However, our data do not support a role of ASIC2 in those facets of mechanoperception.
Figures

Inward currents activated by rapid application of extracellular acid (pH 4.5) recorded from Xenopus oocytes injected with 1 ng of cRNA corresponding to the ASIC2a coding sequence of wild-type or ASIC2 knockout mice. The wild-type ASIC2a sequence was identical to that in the databases (GenBank accession no. AAK40101). In the ASIC2 knockout mice, exon 8 is deleted, leading to a frame-shift before TM2. The carboxy terminus of the targeted ASIC2 protein is 421KAYEVAALLADQREAIRPAWQRRRGREP. The initial pH was 7.4 and the holding potential was −60 mV. Inset, ASIC2a protein cannot be detected in ASIC2 null mice. Western Blot with an antibody directed against the ASIC2a NH2 terminus. ASIC3 COS, ASIC2a COS, 6 μg homogenate from ASIC3 or ASIC2a transfected COS cells, respectively; +/+ and −/−, 20 μg brain homogenate from ASIC2+/+ or ASIC2−/− mice. Neither ASIC2a nor the 7 kDa shorter protein for which the targeted ASIC2 transcript codes are detected in ASIC2−/− mice. The major band of about 70 kDa labelled in brain homogenate from both ASIC2+/+ and ASIC2−/− mice is probably due to a cross-reactivity of the anti-ASIC2a antibody with an unrelated protein. No labelling was obtained in the absence of the anti-ASIC2a antibody (not shown).

A, current traces recorded from an ASIC2+/+ mouse hippocampal neurone. PcTx1 (20 nm ), applied before the drop to pH 5, only partly inhibits the ASIC-like current (left), whereas a coapplication of 300 μm zinc during the drop to pH 6 increases the current amplitude (right). B, current traces recorded from an ASIC2−/− mouse hippocampal neurone. PcTx1 (20 nm ) applied before the drop to pH 5 almost completely inhibits the ASIC-like current (left), whereas a coapplication of 300 μm zinc during the drop to pH 6 induces no increase of the current amplitude (right). C, mean inhibitory effect (IPcTx1/Icontrol) of PcTx1 (20 nm ) on pH 5-induced ASIC-like current recorded from ASIC2+/+ (open bar) and ASIC2−/− mouse hippocampal neurones (filled bar). The number of experiments is shown above the bars. *P < 0.05 compared to ASIC2+/+. D, mean effect (Izinc/Icontrol) of zinc (300 μm ) on pH 6-induced ASIC-like currents recorded from ASIC2+/+ (open bar) and ASIC2−/− mouse hippocampal neurones (filled bar). The number of experiments is shown above the bar. The dashed line indicates the no-effect level. *P < 0.05 compared to ASIC2+/+. The holding potential was −50 mV in all experiments. The zero current level is indicated by a side tick.

Tone bursts (2, 4, 8, 10, 16, 26, 32 and 64 kHz) of increasing intensity (0–100 dB, increment 5 dB) were presented to the ears and compound action potentials (CAP) were recorded from the auditory nerve. The CAP thresholds were defined as the sound pressure level (dB SPL) needed to elicit a measurable response (>5 μV). CAP audiograms shown are from wild-type (black symbols) and ASIC2 knockout mice (shaded symbols). Data for each genotype are means ± s.e.m. (n = 6). The inset shows typical 2.5 kHz low-pass filtered cochlear potentials that were recorded from ASIC2 knockout (shaded trace) and wild-type (black trace) mice evoked by 10 kHz, 40 dB tone bursts. CAP and SP in the inset indicate where the compound action potentials and summating potentials, respectively, were measured.
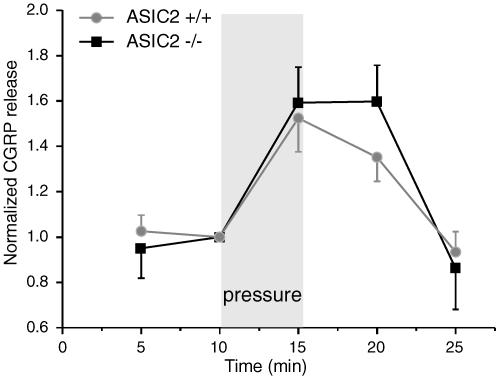
Samples were taken every 5 min and the CGRP content was determined. After 10 min the colon was inflated to 60 mmHg for 5 min. CGRP release was normalized to the control incubation preceding the mechanical stimulation. Values are means ± s.e.m. (n = 5).

Stimulus–response functions of rapidly adapting (RA) fibres to controlled force stimuli during stimulus onset (A) and offset (B). RA fibres with typical dynamic response characteristics respond with high-frequency discharge during stimulus onset (dynamic on response) and fast adaptation without further discharge during the plateau phase of the stimulus. Most of them show a short burst of action potential when the stimulator retracts (dynamic off response). Fibres in both populations similarly code for strength at stimulus onset but not at offset. Values represent means ± s.e.m. of number of action potentials per second (n = 9–12). C, examples of instantaneous discharge frequencies of RA fibres recorded from ASIC2−/− and WT mice stimulated with 100 and 150 mN. Upward and downward arrows indicate stimulus onset and relief, respectively. The ASIC2−/− fibre shown had a short dynamic off response when the stimulus was removed. The insets show the unitary action potentials with the thin lines depicting one standard deviation as a measure for noise levels.
References
-
- Akopian AN, Chen CC, Ding Y, Cesare P, Wood JN. A new member of the acid-sensing ion channel family. Neuroreport. 2000;11:2217–2222. - PubMed
-
- Alessandri-Haber N, Yeh JJ, Boyd AE, Parada CA, Chen X, Reichling DB, et al. Hypotonicity induces TRPV4-mediated nociception in rat. Neuron. 2003;39:497–511. - PubMed
-
- Babinski K, Catarsi S, Biagini G, Seguela P. Mammalian ASIC2a and ASIC3 subunits co-assemble into heteromeric proton-gated channels sensitive to Gd3+ J Biol Chem. 2000;275:28519–28525. - PubMed
-
- Babinski K, Le KT, Seguela P. Molecular cloning and regional distribution of a human proton receptor subunit with biphasic functional properties. J Neurochem. 1999;72:51–57. - PubMed
-
- Baron A, Schaefer L, Lingueglia E, Champigny G, Lazdunski M. Zn2+ and H+ are coactivators of acid-sensing ion channels. J Biol Chem. 2001;276:35361–35367. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases