

Immune responses in healthy and allergic individuals are characterized by a fine balance between allergen-specific T regulatory 1 and T helper 2 cells
- PMID: 15173208
- PMCID: PMC2211782
- DOI: 10.1084/jem.20032058
Immune responses in healthy and allergic individuals are characterized by a fine balance between allergen-specific T regulatory 1 and T helper 2 cells
Abstract
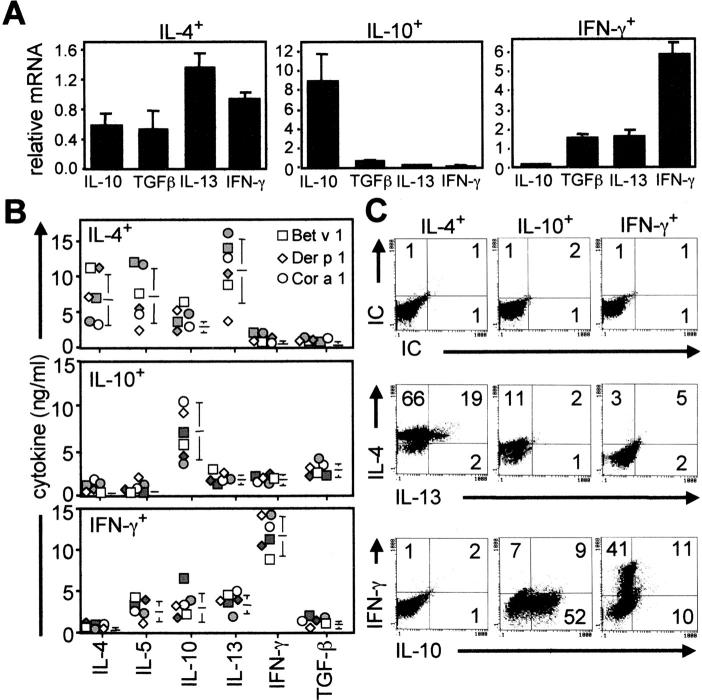
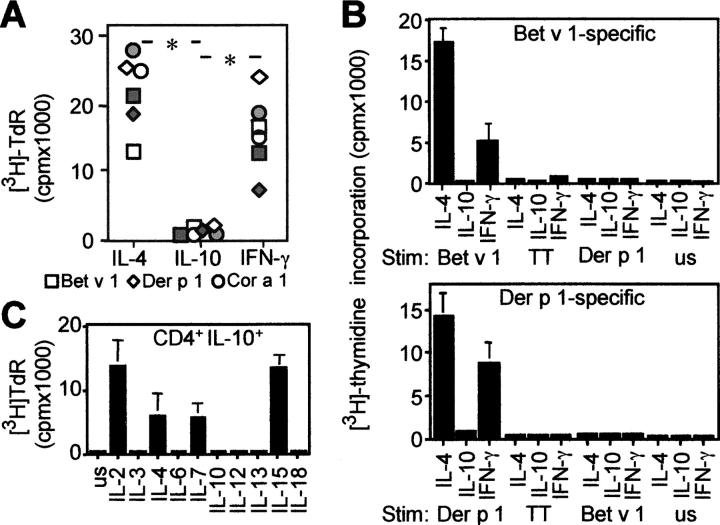
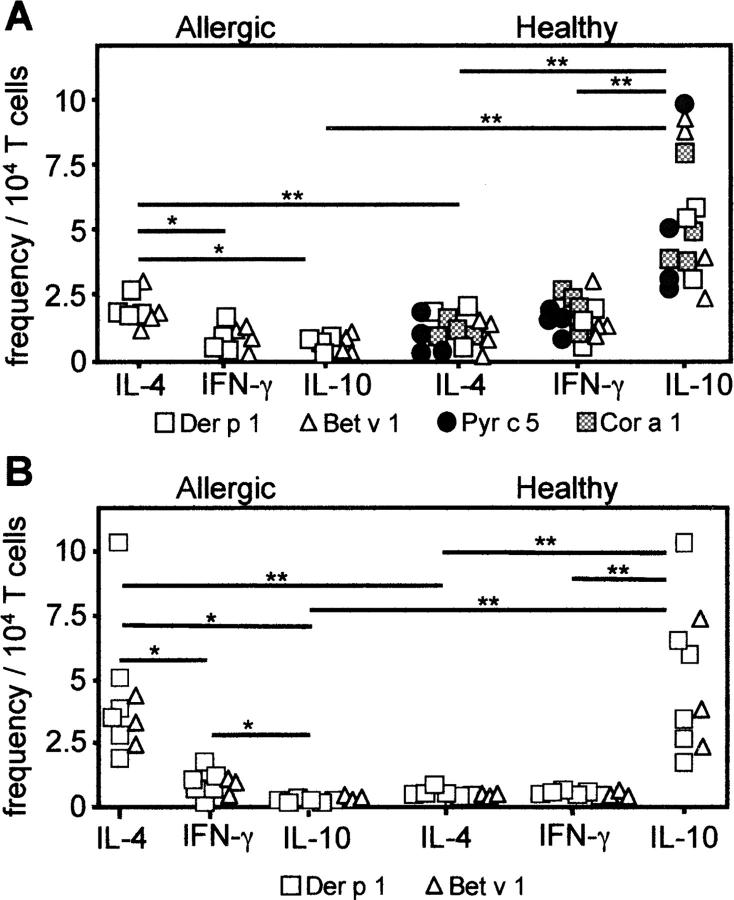
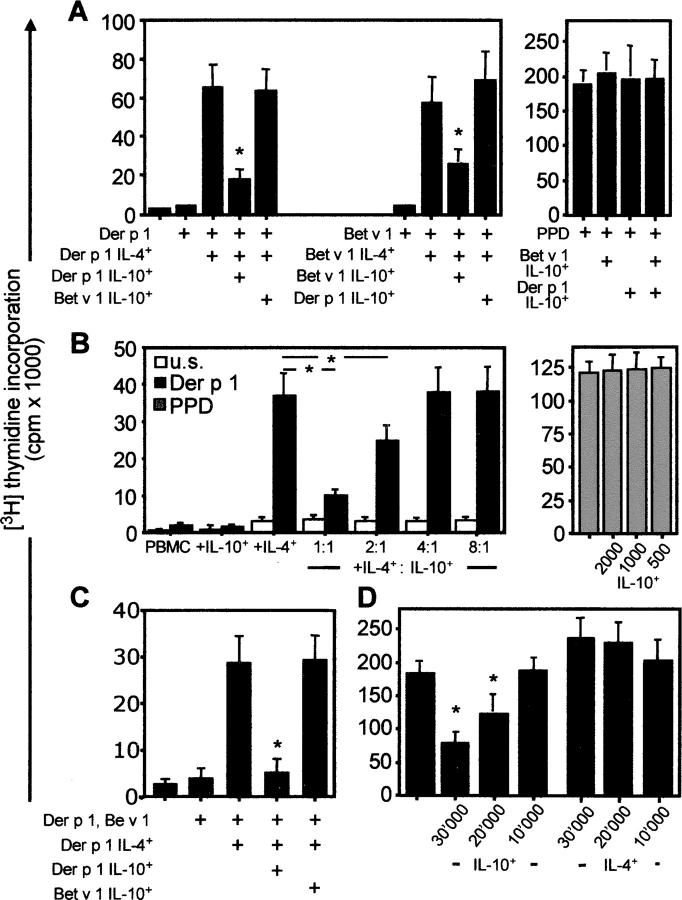
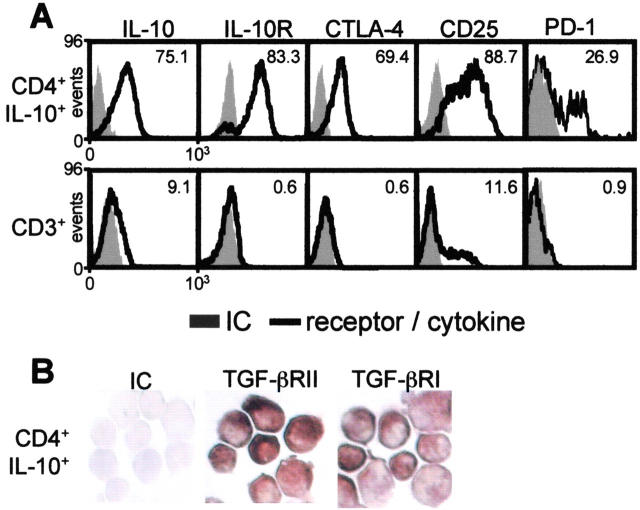
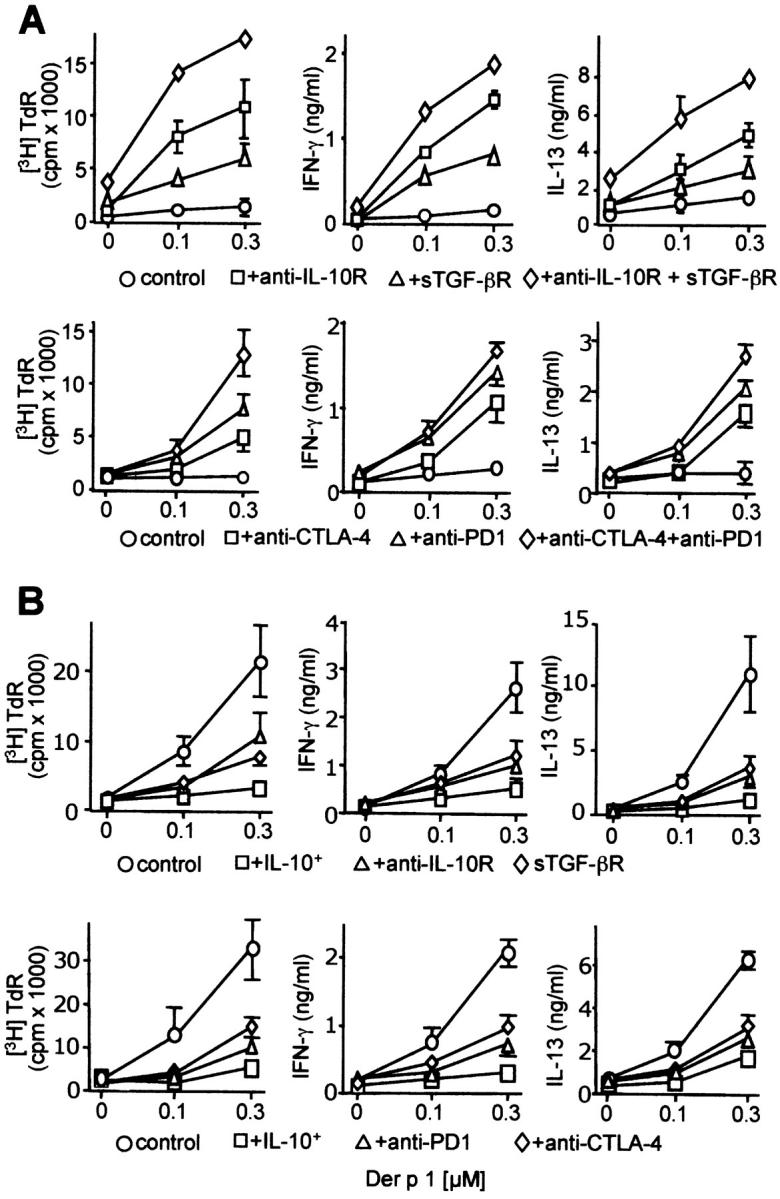
The mechanisms by which immune responses to nonpathogenic environmental antigens lead to either allergy or nonharmful immunity are unknown. Single allergen-specific T cells constitute a very small fraction of the whole CD4+ T cell repertoire and can be isolated from the peripheral blood of humans according to their cytokine profile. Freshly purified interferon-gamma-, interleukin (IL)-4-, and IL-10-producing allergen-specific CD4+ T cells display characteristics of T helper cell (Th)1-, Th2-, and T regulatory (Tr)1-like cells, respectively. Tr1 cells consistently represent the dominant subset specific for common environmental allergens in healthy individuals; in contrast, there is a high frequency of allergen-specific IL-4-secreting T cells in allergic individuals. Tr1 cells use multiple suppressive mechanisms, IL-10 and TGF-beta as secreted cytokines, and cytotoxic T lymphocyte antigen 4 and programmed death 1 as surface molecules. Healthy and allergic individuals exhibit all three allergen-specific subsets in different proportions, indicating that a change in the dominant subset may lead to allergy development or recovery. Accordingly, blocking the suppressor activity of Tr1 cells or increasing Th2 cell frequency enhances allergen-specific Th2 cell activation ex vivo. These results indicate that the balance between allergen-specific Tr1 cells and Th2 cells may be decisive in the development of allergy.
Figures
References
-
- Teale, J.M., and N.R. Klinman. 1980. Tolerance as an active process. Nature. 288:385–387. - PubMed
-
- Parijs, L.V., and A.K. Abbas. 1998. Homeostasis and self-tolerance in the immune system: turning lymphocytes off. Science. 280:243–247. - PubMed
-
- Mosmann, T.R., H. Cherwinski, M.W. Bond, M.A. Giedlin, and R.L. Coffman. 1986. Two types of murine helper T cell clones. 1. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 136:2348–2357. - PubMed
-
- Romagnani, S. 1994. Lymphokine production by human T cells in disease states. Annu. Rev. Immunol. 12:227–257. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
