

The peroxisome deficient Arabidopsis mutant sse1 exhibits impaired fatty acid synthesis
- PMID: 15173561
- PMCID: PMC514117
- DOI: 10.1104/pp.103.036772
The peroxisome deficient Arabidopsis mutant sse1 exhibits impaired fatty acid synthesis
Abstract
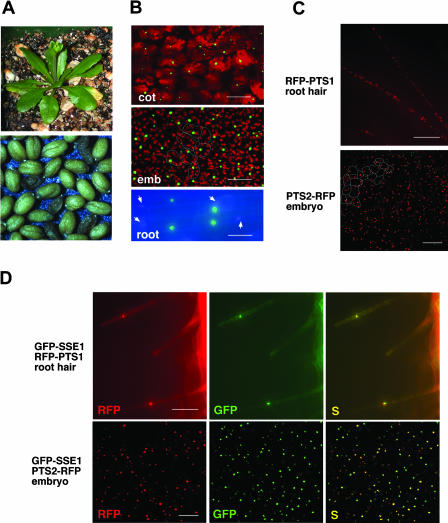
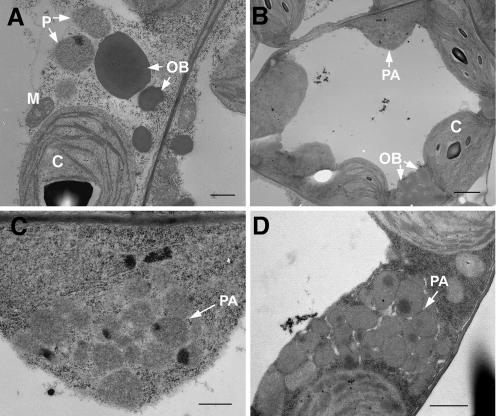
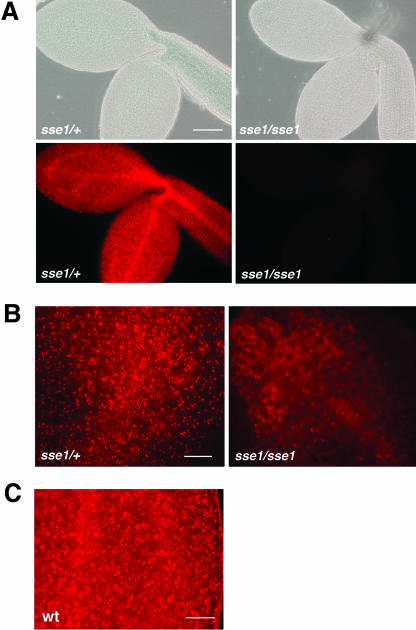
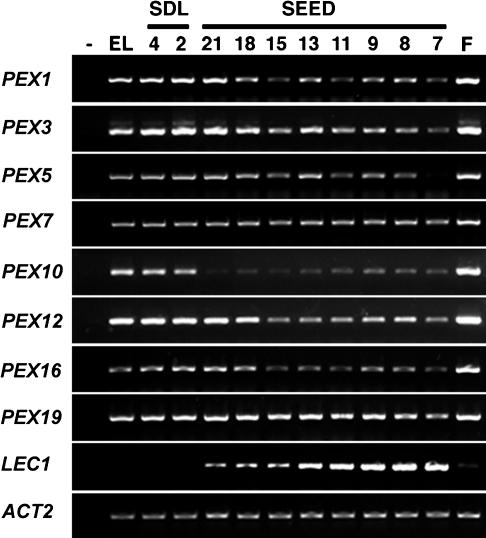
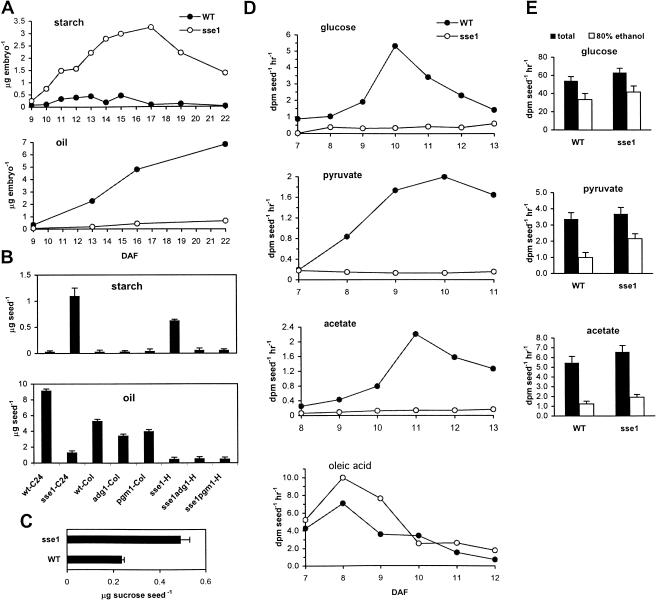
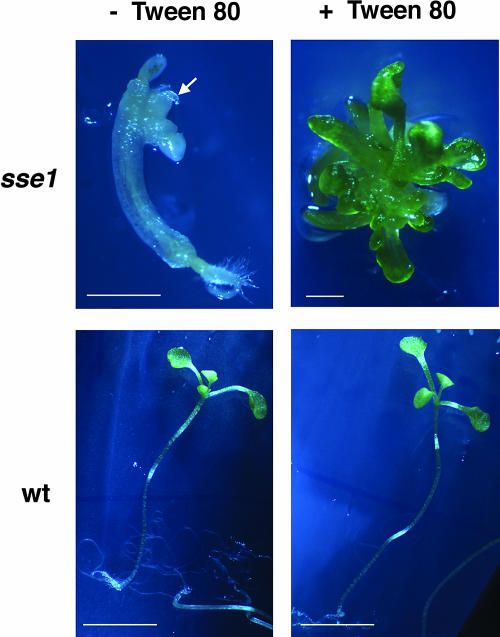
The Arabidopsis Shrunken Seed 1 (SSE1) gene encodes a homolog of the peroxisome biogenesis factor Pex16p, and a loss-of-function mutation in this gene alters seed storage composition. Two lines of evidence support a function for SSE1 in peroxisome biogenesis: the peroxisomal localization of a green fluorescent protein-SSE1 fusion protein and the lack of normal peroxisomes in sse1 mutant embryos. The green fluorescent protein-SSE1 colocalizes with the red fluorescent protein (RFP)-labeled peroxisomal markers RFP-peroxisome targeting signal 1 and peroxisome targeting signal 2-RFP in transgenic Arabidopsis. Each peroxisomal marker exhibits a normal punctate peroxisomal distribution in the wild type but not the sse1 mutant embryos. Further studies reported here were designed toward understanding carbon metabolism in the sse1 mutant. A time course study of dissected embryos revealed a dramatic rate decrease in oil accumulation and an increase in starch accumulation. Introduction of starch synthesis mutations into the sse1 background did not restore oil biosynthesis. This finding demonstrated that reduction in oil content in sse1 is not caused by increased carbon flow to starch. To identify the blocked steps in the sse1 oil deposition pathway, developing sse1 seeds were supplied radiolabeled oil synthesis precursors. The ability of sse1 to incorporate oleic acid, but not pyruvate or acetate, into triacylglycerol indicated a defect in the fatty acid biosynthetic pathway in this mutant. Taken together, the results point to a possible role for peroxisomes in the net synthesis of fatty acids in addition to their established function in lipid catabolism. Other possible interpretations of the results are discussed.
Figures

References
-
- An YQ, McDowell JM, Huang S, Mckinney EC, Chambliss S, Meagher RB (1996) Strong, constitutive expression of the Arabidopis ACT2/ACT8 actin subclass in vegetative tissues. Plant J 10: 107–121 - PubMed
-
- Arai Y, Nakashita H, Suzuki Y, Kobayashi Y, Shimizu T, Yasuda M, Doi Y, Yamaguchi I (2002) Synthesis of a novel class of polyhydroxyalkanoates in peroxisomes, and their use in monitoring short-chain-length intermediates of β-oxidation. Plant Cell Physiol 43: 555–562 - PubMed
-
- Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J, Ruvkun G (2003) Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 421: 268–272 - PubMed
-
- Buchanan BB, Gruissem W, Jones RL (2000) Biochemistry and Molecular Biology of Plants. American Society of Plant Physiologists, Rockville, Maryland
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
