

Salicylate accumulation inhibits growth at chilling temperature in Arabidopsis
- PMID: 15173571
- PMCID: PMC514138
- DOI: 10.1104/pp.104.041293
Salicylate accumulation inhibits growth at chilling temperature in Arabidopsis
Abstract
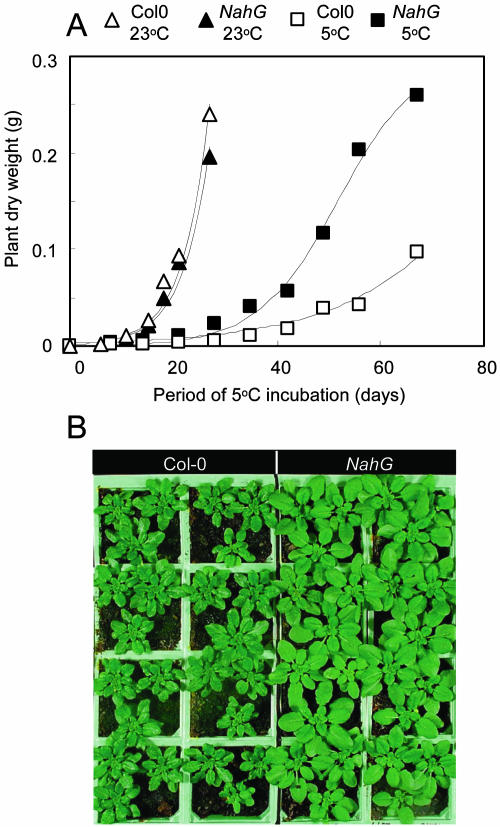
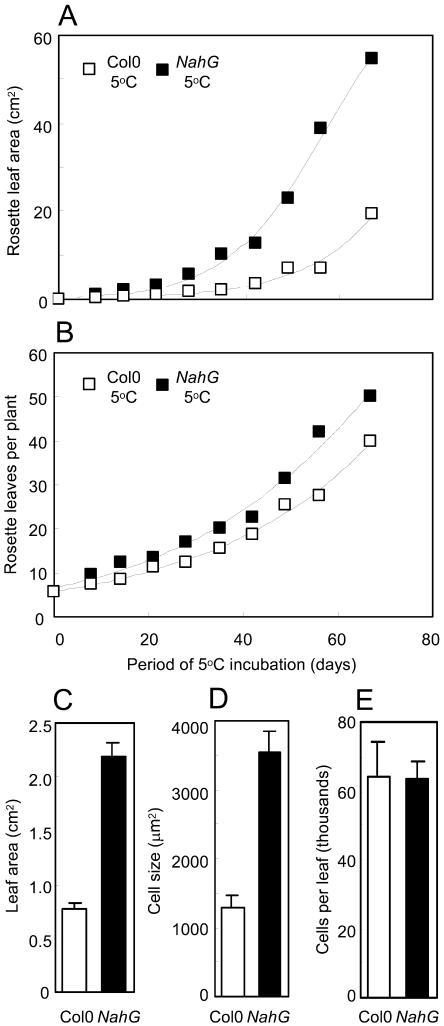
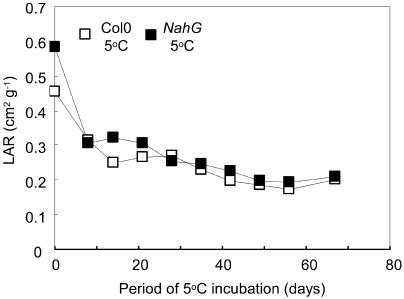
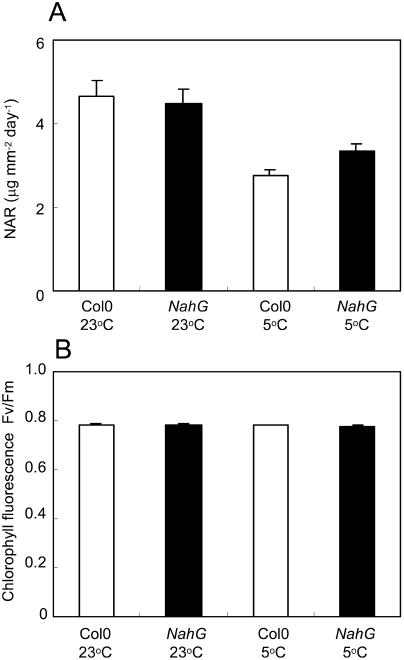
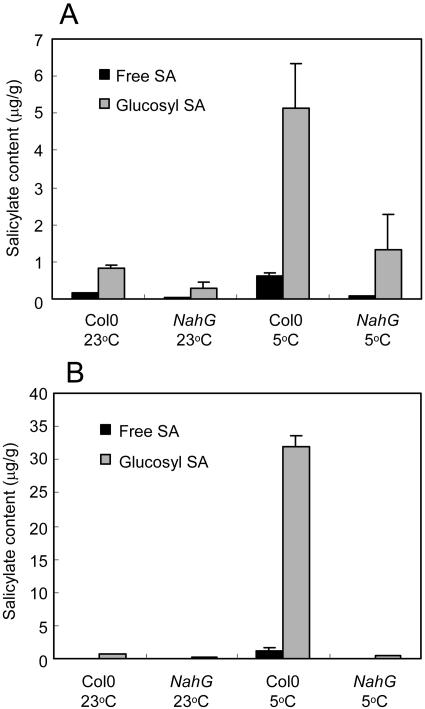
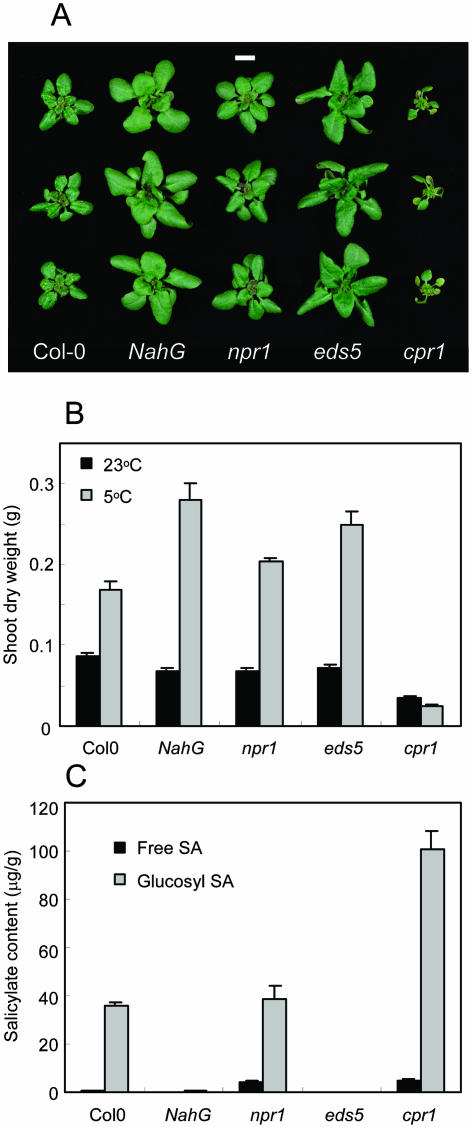
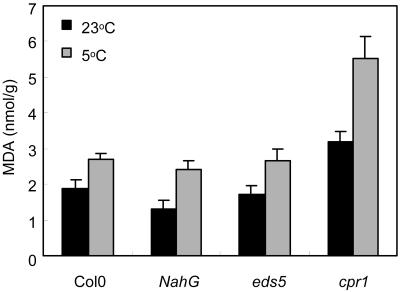
The growth of Arabidopsis plants in chilling conditions could be related to their levels of salicylic acid (SA). Plants with the SA hydroxylase NahG transgene grew at similar rates to Col-0 wild types at 23 degrees C, and growth of both genotypes was slowed by transfer to 5 degrees C. However, at 5 degrees C, NahG plants displayed relative growth rates about one-third greater than Col-0, so that by 2 months NahG plants were typically 2.7-fold larger. This resulted primarily from greater cell expansion in NahG rosette leaves. Specific leaf areas and leaf area ratios remained similar in both genotypes. Net assimilation rates were similar in both genotypes at 23 degrees C, but higher in NahG at 5 degrees C. Chlorophyll fluorescence measurements revealed no PSII photodamage in chilled leaves of either genotype. Col-0 shoots at 5 degrees C accumulated SA, particularly in glucosylated form. SA in NahG shoots showed similar tendencies at 5 degrees C, but at greatly depleted levels. Catechol was not detected as a metabolite of the NahG transgene product. We also examined growth and SA levels in SA signaling and metabolism mutants at 5 degrees C. The partially SA-insensitive npr1 mutant displayed growth intermediate between NahG and Col-0, while the SA-deficient eds5 mutant behaved like NahG. In contrast, the cpr1 mutant at 5 degrees C accumulated very high levels of SA and its growth was much more inhibited than wild type. At both temperatures, cpr1 was the only SA-responsive genotype in which oxidative damage (measured as thiobarbituric acid-reactive substances) was significantly different from wild type.
Figures
References
-
- Allen DJ, Ort DR (2001) Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci 6: 36–42 - PubMed
-
- Bernard F, Shaker-Bazarnov H, Kaviani B (2002) Effects of salicylic acid on cold preservation and cryopreservation of encapsulated embryonic axes of Persian lilac (Melia azedarach L.). Euphytica 123: 85–88
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
