

Leukocyte engagement of fibrin(ogen) via the integrin receptor alphaMbeta2/Mac-1 is critical for host inflammatory response in vivo
- PMID: 15173886
- PMCID: PMC419487
- DOI: 10.1172/JCI20741
Leukocyte engagement of fibrin(ogen) via the integrin receptor alphaMbeta2/Mac-1 is critical for host inflammatory response in vivo
Abstract
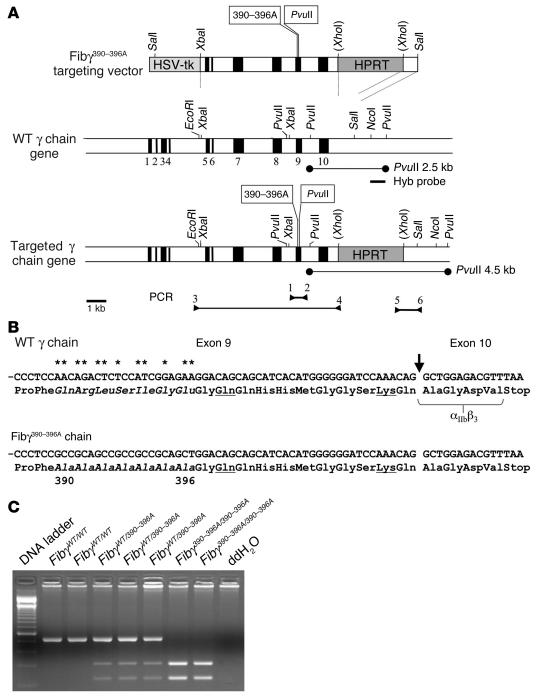
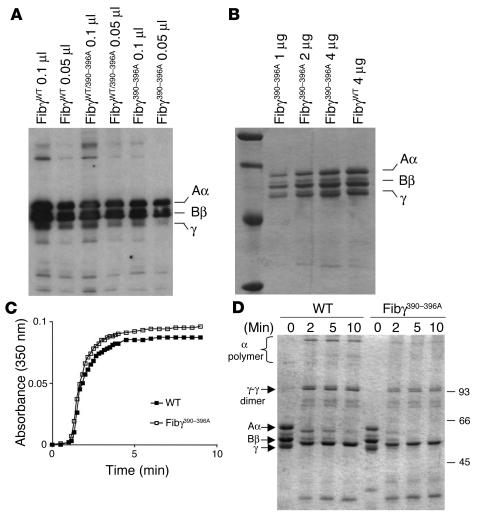
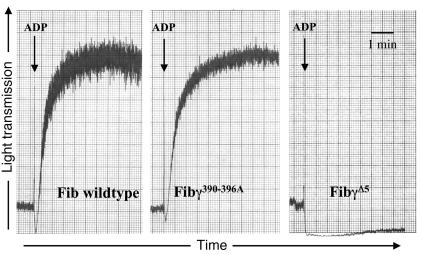
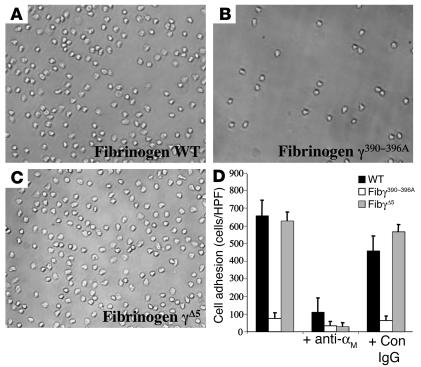
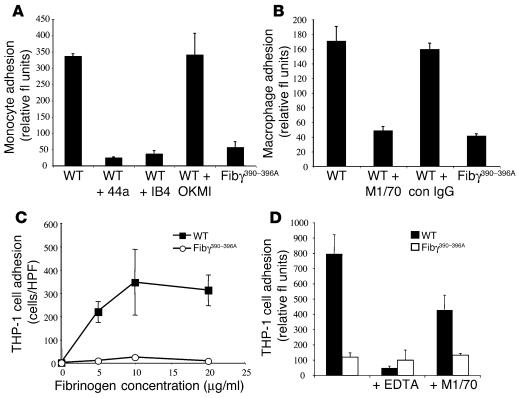
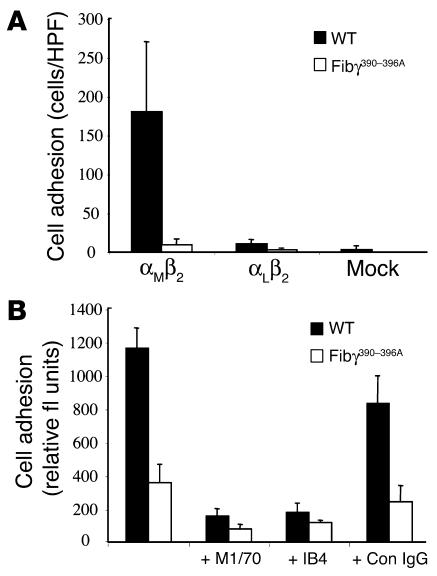
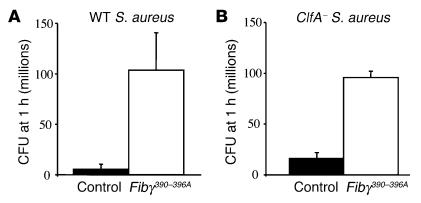
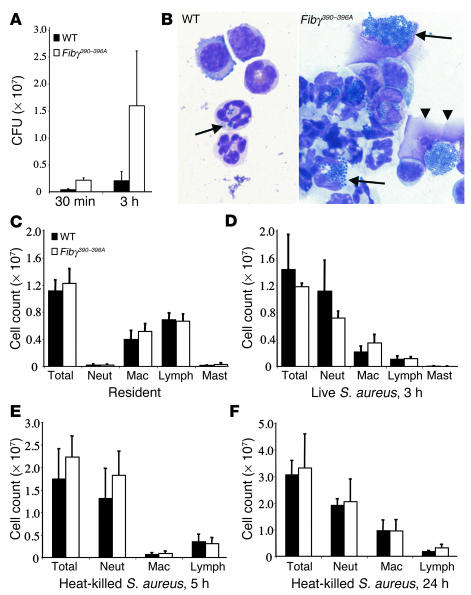
The leukocyte integrin alpha(M)beta(2)/Mac-1 appears to support the inflammatory response through multiple ligands, but local engagement of fibrin(ogen) may be particularly important for leukocyte function. To define the biological significance of fibrin(ogen)-alpha(M)beta(2) interaction in vivo, gene-targeted mice were generated in which the alpha(M)beta(2)-binding motif within the fibrinogen gamma chain (N(390)RLSIGE(396)) was converted to a series of alanine residues. Mice carrying the Fibgamma(390-396A) allele maintained normal levels of fibrinogen, retained normal clotting function, supported platelet aggregation, and never developed spontaneous hemorrhagic events. However, the mutant fibrinogen failed to support alpha(M)beta(2)-mediated adhesion of primary neutrophils, macrophages, and alpha(M)beta(2)-expressing cell lines. The elimination of the alpha(M)beta(2)-binding motif on fibrin(ogen) severely compromised the inflammatory response in vivo as evidenced by a dramatic impediment in leukocyte clearance of Staphylococcus aureus inoculated into the peritoneal cavity. This defect in bacterial clearance was due not to diminished leukocyte trafficking but rather to a failure to fully implement antimicrobial functions. These studies definitively demonstrate that fibrin(ogen) is a physiologically relevant ligand for alpha(M)beta(2), integrin engagement of fibrin(ogen) is critical to leukocyte function and innate immunity in vivo, and the biological importance of fibrinogen in regulating the inflammatory response can be appreciated outside of any alteration in clotting function.
Figures
Similar articles
-
Fibrin(ogen)-alpha M beta 2 interactions regulate leukocyte function and innate immunity in vivo.Exp Biol Med (Maywood). 2004 Dec;229(11):1105-10. doi: 10.1177/153537020422901104. Exp Biol Med (Maywood). 2004. PMID: 15564436 Review.
-
Fibrin(ogen) drives repair after acetaminophen-induced liver injury via leukocyte αMβ2 integrin-dependent upregulation of Mmp12.J Hepatol. 2017 Apr;66(4):787-797. doi: 10.1016/j.jhep.2016.12.004. Epub 2016 Dec 10. J Hepatol. 2017. PMID: 27965156 Free PMC article.
-
Fibrin(ogen) exacerbates inflammatory joint disease through a mechanism linked to the integrin alphaMbeta2 binding motif.J Clin Invest. 2007 Nov;117(11):3224-35. doi: 10.1172/JCI30134. J Clin Invest. 2007. PMID: 17932565 Free PMC article.
-
Amelioration of Duchenne muscular dystrophy in mdx mice by elimination of matrix-associated fibrin-driven inflammation coupled to the αMβ2 leukocyte integrin receptor.Hum Mol Genet. 2012 May 1;21(9):1989-2004. doi: 10.1093/hmg/dds012. Epub 2012 Mar 1. Hum Mol Genet. 2012. PMID: 22381526 Free PMC article.
-
Targeting the coagulation factor fibrinogen for arthritis therapy.Curr Pharm Biotechnol. 2011 Sep;12(9):1497-506. doi: 10.2174/138920111798281144. Curr Pharm Biotechnol. 2011. PMID: 21401516 Review.
Cited by
-
Fibrinogen, Fibrin, and Fibrin Degradation Products in COVID-19.Curr Drug Targets. 2022;23(17):1593-1602. doi: 10.2174/1389450123666220826162900. Curr Drug Targets. 2022. PMID: 36029073 Free PMC article. Review.
-
The intersection of coagulation activation and inflammation after injury: What you need to know.J Trauma Acute Care Surg. 2024 Mar 1;96(3):347-356. doi: 10.1097/TA.0000000000004190. Epub 2023 Nov 13. J Trauma Acute Care Surg. 2024. PMID: 37962222 Free PMC article. No abstract available.
-
Aluminum adjuvants elicit fibrin-dependent extracellular traps in vivo.Blood. 2010 Dec 9;116(24):5191-9. doi: 10.1182/blood-2010-03-275529. Epub 2010 Sep 27. Blood. 2010. PMID: 20876456 Free PMC article.
-
Integrin alphaDbeta2, an adhesion receptor up-regulated on macrophage foam cells, exhibits multiligand-binding properties.Blood. 2006 Feb 15;107(4):1643-50. doi: 10.1182/blood-2005-06-2509. Epub 2005 Oct 20. Blood. 2006. PMID: 16239428 Free PMC article.
-
Platelet-derived TLT-1 is a prognostic indicator in ALI/ARDS and prevents tissue damage in the lungs in a mouse model.Blood. 2018 Dec 6;132(23):2495-2505. doi: 10.1182/blood-2018-03-841593. Epub 2018 Oct 3. Blood. 2018. PMID: 30282800 Free PMC article.
References
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76:301–314. - PubMed
-
- Anderson, D.C., Kishimoto, T.K., and Smith, C.W. 1995. Leukocyte adhesion deficiency and other disorders of leukocyte adherence and motility. In The metabolic and molecular bases of inherited disease. W.S. Sly, C.R. Scriver, A.L. Beaudet, and D. Valle, editors. McGraw-Hill. New York, New York, USA. 3955–3994.
-
- Larson RS, Springer TA. Structure and function of leukocyte integrins. Immunol. Rev. 1990;114:181–217. - PubMed
-
- Van der Vieren M, et al. A novel leukointegrin, αdβ2, binds preferentially to ICAM-3. Immunity. 1995;3:683–690. - PubMed
-
- Coxon A, et al. A novel role for the β2 integrin CD11b/CD18 in neutrophil apoptosis: a homeostatic mechanism in inflammation. Immunity. 1996;5:653–666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
